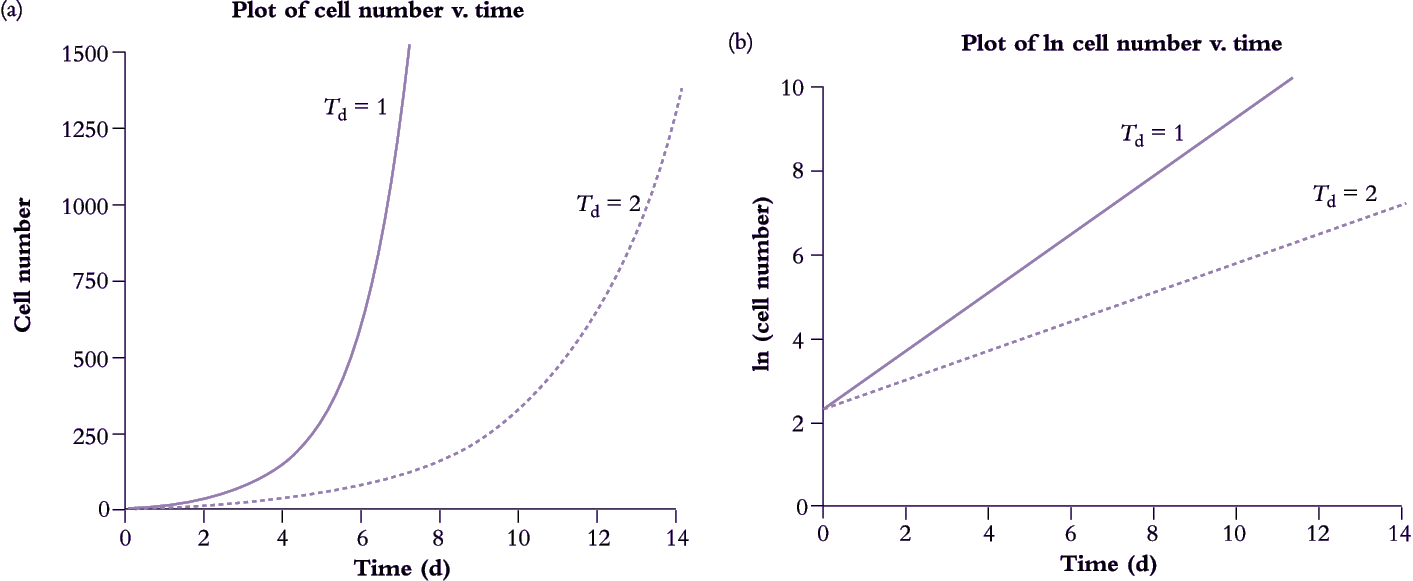
In whole plants, cell number is an impractical measure of growth. Instead, fresh or oven-dried biomass (W) is generally taken as a surrogate for cell growth and referenced to the number of days elapsed between successive observations. Relative growth rate is now known as RGR rather than r and measured in days or weeks rather than hours.
Relative growth rate, RGR (d–1), can be expressed in terms of differential calculus as \(RGR=\frac{1}{W}\frac{\text{d}W}{\text{d}t}\) (compare Equation 6.2.) so that RGR is increment in dry mass (dW) per increment in time (dt) divided by existing biomass (W). Averaged over a time interval t1 to t2 during which time biomass increases from W1 to W2, RGR (d–1) can be calculated from
\[\text{RGR}=\frac{\text{ln }W_2-\text{ln } W_1}{t_2-t_1} \tag{6.5} \]
which is analogous to Eq, 6.4. Net gain in biomass (W) is the outcome of CO2 assimilation by leaves minus respiratory loss by the entire plant. Leaf area can therefore be viewed as a driving variable, and biomass increment (dW) per unit time (dt) can then be divided by leaf area (A) to yield the net assimilation rate, NAR (g m–2 d–1), where
\[\text{NAR}=\frac{1}{A}\frac{\text{d}W}{\text{d}T} \tag{6.6}\]
Averaged over a short time interval (t1 to t2 days) and provided whole-plant biomass and leaf area are linearly related (see Radford 1967),
\[\text{NAR}=\left(\frac{W_2-W_1}{t_2-t_1}\right)\left(\frac{\text{ln }A_2-\text{ln }A_1}{A_2-A_1}\right) \tag{6.7}\]
NAR thus represents a plant’s net photosynthetic effectiveness in capturing light, assimilating CO2 and storing photoassimilate. Variation in NAR can derive from differences in canopy architecture and light interception, photosynthetic activity of leaves, respiration, transport of photoassimilate and storage capacity of sinks, or even the chemical nature of stored products.
The following treatment assumes for simplicity that photosynthesis and the assimilation of CO2 occurs only in leaves, even though for many herbaceous or succulent species it occurs to a lesser degree also in stems. Since leaf area is a driving variable for whole-plant growth, the proportion of plant biomass invested in leaf area will have an important bearing on RGR, and can be conveniently defined as leaf area ratio, LAR (m2 g–1), where
\[\text{LAR}=\frac{A}{W}\tag{6.8}\]
LAR can be factored into two components: specific leaf area (SLA) and leaf weight ratio (LWR). SLA is the ratio of leaf area (A) to leaf mass (WL) (m2 g–1) and LWR is the ratio of leaf mass (WL) to total plant mass (W) (dimensionless). Thus,
\[\begin{align} \text{LAR} &= \frac{A}{W_L}\frac{W_L}{W} \\
&= \text{SLA} \times \text{LWR} \end{align} \tag{6.9} \]
As an aside, average LAR over the growth interval t1 to t2 is
\[\text{LAR}=\frac{1}{2} \left( \frac{A_1}{W_1}+\frac{A_2}{W_2} \right) \tag{6.10}\]
Expressed this way, LAR becomes a more meaningful growth index than A/W (Equation 6.8) and can help resolve sources of variation in RGR.
If both A and W are increasing exponentially so that W is proportional to A, it follows that
\[\frac{1}{W}\frac{\text{d}W}{\text{d}t}=\frac{1}{A}\frac{\text{d}W}{\text{d}t}\times\frac{A}{W}\tag{6.11}\]
0r (substituting Equations 6.5, 6.6 and 6.8)
\[\text{RGR}=\text{NAR}\times \text{LAR} \tag{6.12}\]
As LAR can be broken into SLA and LWR (Equation 6.9) then
\[\text{RGR}=\text{NAR}\times \text{LWR} \times \text{SLA} \tag{6.13}\]
Sources of variation in RGR partitioned this way provide useful insights on driving variables in process physiology and ecology. For an expanded discussion on methodological issues associated with the determination of RGR in experimental populations see Poorter and Lewis (1986).
Increases in leaf area over time can be a more useful basis for measuring plant growth rates than biomass increases, particularly as non-destructive techniques for measuring leaf area are now available. Plant growth rate can be measured as the relative increase in leaf area over time, by substituting total plant leaf area for total biomass in the conventional RGR equation.
\[\text{RGR}_\text{A} =\frac{\text{ln } LA_2 - \text{ln } LA_1}{t_2-t_1} \tag{6.14} \]
where RGRA is relative leaf area expansion rate, LA is total plant leaf area and t is time at two time intervals, t1 and t2, preferably 2-3 days apart.
Growth indices in summary
Five key indices are commonly derived as an aid to understanding growth responses. Mathematical and functional definitions of those terms are summarised below.
|
Growth index
|
Mathematical definition
|
Units
|
Functional definition
|
|
Relative growth rate
RGR
|
\(\frac{1}{W}\frac{\text{d}W}{\text{d}t}\)
|
d-1
|
Rate of mass increase per unit mass present (efficiency of growth with respect to biomass)
|
|
Net assimilation rate
NAR
|
\(\frac{1}{A}\frac{\text{d}W}{\text{d}t}\)
|
g m-2 d-1
|
Rate of mass increase per unit leaf area (efficiency of leaves in generating biomass)
|
|
Leaf area ratio
LAR
|
\(\frac{A}{W}\)
|
m2 g-1
|
Ratio of leaf area to total plant mass (a measure of ‘leafiness’ or photosynthetic area relative to respiratory mass)
|
|
Specific leaf area
SLA
|
\(\frac{A}{W_L}\)
|
m2 g-1
|
Ratio of leaf area to leaf mass (a measure of thickness of leaves relative to area)
|
|
Leaf weight ratio
LWR
|
\(\frac{W_L}{W}\)
|
dimension-less
|
Ratio of leaf mass to total plant mass (a measure of biomass allocation to leaves)
|