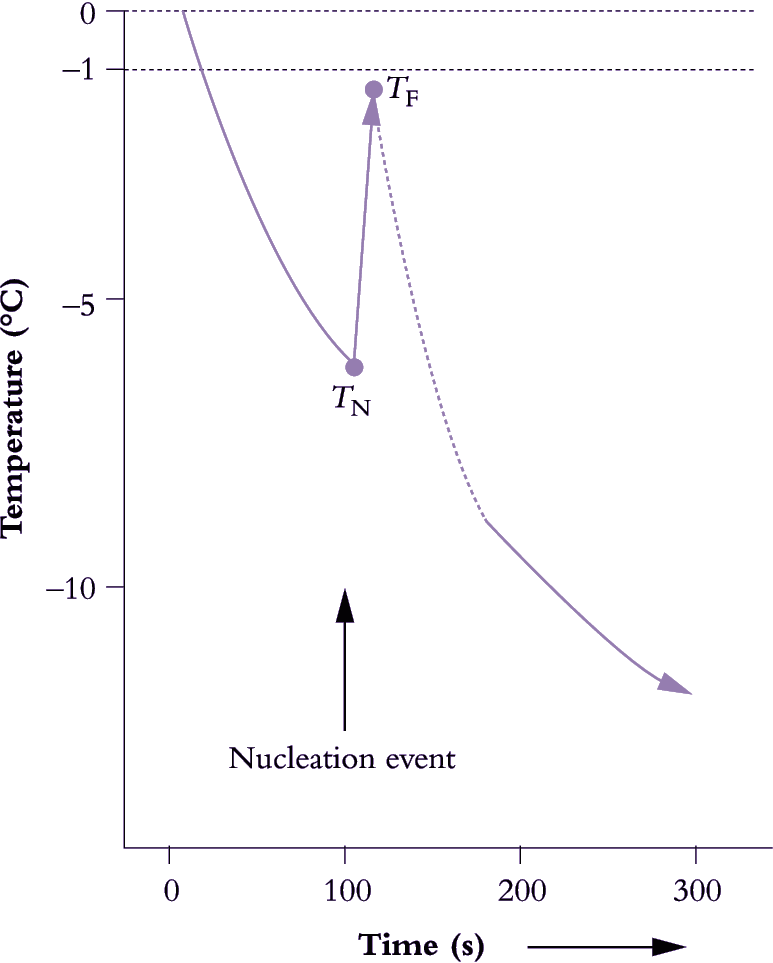
During a frost episode in nature, plants experience a sequence of events similar to that summarised in Figure 14.25. Tissue water supercools, and cell sap freezing point is depressed by osmotically active material. Moreover, plants are also equipped with ice nucleating agents that can be of either plant or bacterial origin (e.g. Pseudomonas syringae). As with solutions, formation of ice in plants is accompanied by a release of heat. This exothermic response can be detected by sensitive infrared thermography, and has been used to trace ice propagation during a freezing event in leaves and shoots (Wisniewski et al. 1997).
When plants experience a frost, ice initially forms within intercellular spaces (apoplasm) where solute concentration is least. Water potential in that region is immediately lowered and water molecules migrate from symplasm to apoplasm across plasmalemma membranes and towards regions of ice crystallisation. Water potential in the apoplasm will decrease by about 1.2 MPa per degree below 0°C, so that an apoplasm at –4°C will have a water potential of around –4.8 MPa (i.e. equivalent to an osmotic pressure twice that of seawater), which will dehydrate the symplasm. As one positive trade-off of the high solute concentration, the symplasm is less likely to freeze. In addition, plasmalemma membranes discourage entry of ice crystals that might otherwise seed ice crystal formation within the symplasm. Nevertheless, partial dehydration does perturb cell biochemistry due to concentration of metabolites, and the accompanying shrinkage of cells and organelles generates structural tensions.
In frost-sensitive material, cell disruption follows the course outlined above, which unfolds over about 3°C on a cooling curve. Membrane integrity might be lost at around –4°C as apoplasm ice formation ruptures membranes. Intracellular freezing ensues at around –7°C and is inevitably lethal due to the combined effects of membrane injury, symplasm dehydration and protein denaturation.
Tissue damage following freezing can be demonstrated by a loss of membrane integrity, metabolite leakage and failure to achieve either plasmolysis or deplasmolysis. Solutions bathing frozen/thawed tissue thus show a sudden increase in electrical conductivity according to freezing damage, and that value then serves as a reliable assay for comparative frost tolerance. For example, population screening based on metabolite leakage has enabled breeding for improved frost tolerance in Eucalyptus nitens for plantation forestry (Raymond et al. 1992).
Leaves on temperate plants often need to accommodate ice formation, and Rhododendron provides such an example. Frozen leaves appear wilted, but regain their normal turgid appearance following thawing. Camellia leaves behave similarly. They take on a semi-transparent appearance when frozen, but recover without damage when thawed. In both cases, their altered physical appearance at low temperature is due to formation of a frostblaze, that is, a lens of ice crystals that forms between layers of tissue that are readily cleaved. Ice localised in this way is rendered harmless, and is lost easily on thawing.
Frost hardiness is a dynamic and composite property of plant cells involving cell size, wall thickness, osmotic pressure of cell sap and membrane properties, all of which can feature in either delaying onset or diminishing adverse consequences of ice formation (Steponkus et al. 1993). In any plant, organs that are growing rapidly are frost sensitive, and this is especially the case with early spring growth. Frost hardiness is thus least during the growing season, but increases during autumn and reaches a peak in winter with acclimation to low temperatures. This is an adaptive feature of perennial plants that is attuned to seasonal necessity. Moreover, the extent of this hardening process is heritable, and requires low temperature for onset and maintenance. Return to milder conditions can lead to dehardening with disastrous consequences for horticulture when an early spring temperature increase is followed by an unseasonal period of cold nights. Frost prevention then becomes crucial for reducing damage to buds, flowers or fruitlets. Susceptibility to low temperatures is dependent on stage of morphological development of plant organs. As spring temperatures increase, fruit develop into buds, flowers then fruitlets with a concomitant increase in susceptibility to cold temperatures. In many fruit trees, buds emerging from dormancy can tolerate temperatures as low as -16 to -17oC, while at full bloom critical temperatures for 10% kill of buds after 30 minutes of exposure -2 to -3oC are -1.5 to 3.0oC. To prevent frost damage, growers generally commence frost protection measures at least 10C above the 10% critical temperature