Marilyn Ball, Research School of Biology, Australian National University
Re-establishment of eucalypts in open pastures of the New South Wales tablelands is problematic, with up to 80% of young trees failing to survive beyond three years. An urgent need for landscape restoration following deforestatation and overgrazing of these areas has become widely recognised, and a successful strategy for tree establishment is crucial to that process.
Photoinhibition of young trees at low temperature was soon recognised as a factor in those early losses (Ball et al. 1991). The first clue that sunlight × temperature was an issue came from two chance observations of unusual patterns in seedling establishment.
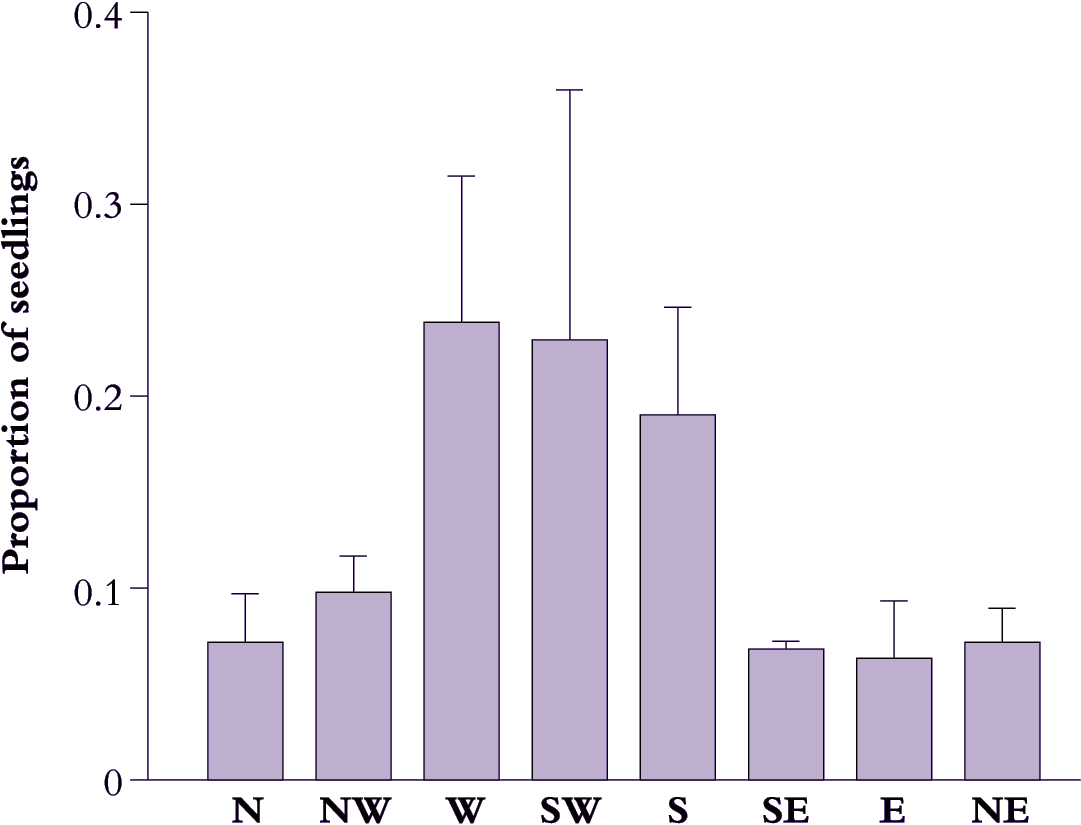
The first observation concerns mountain beech (Nothofagus solandri). This is a dominant canopy tree in subalpine forests of New Zealand, and patterns of seedling regeneration around isolated trees on the Waimakariri floodplain are highly characteristic. While 95% of young seedlings occur beneath the canopy of a parent tree, they are not distributed randomly. Rather, they tend to cluster on the western side, and 65% occur in a narrow sector between 165°S and 285°S (Figure 2). Wind effects on seed dispersal are not responsible because prevailing winds blow in the opposite direction; however, seedlings on the south to western sides of parent trees are protected from direct sunlight all day throughout winter, and from morning sun in spring and early summer.
The second observation concerns a similar asymmetric pattern of regenerating seedlings the occurs under isolated trees of snow gum (Eucalyptus pauciflora) growing along the Orroral Valley in southeastern Australia. In this case, tiny seedlings probably establish randomly around a parent tree during favourable seasons, but are subsequently culled by adverse conditions to produce this now familiar pattern of seedling regeneration.
In both cases, the combination of intense cold and strong sunlight seemed to be proving adverse to seedling survival because the asymmetry in seedling establishment roughly coincided with morning shadows cast by parent trees. Chris Holly tested this idea with artifical shelters on Eucalyptus polyanthemos seedlings in an open pasture (Holly et al. 1994). Seedlings were planted either on open ground or in individual shelters consisting of open-topped cylinders made from one of three types of shadecloth transmitting 30%, 50% or 70% sunlight. Air temperatures inside and outside of these shelters differed by less than 2°C so that irradiance was the major factor that differed between treatments. During winter, leaves became photoinhibited as indicated by a loss in variable fluorescence (Fv/Fm from in vivo chlorophyll a fluorescence measured in situ; Section 1.2.5). The extent of this loss in variable fluorescence (measured pre-dawn) was in direct proportion to treatment irradiance. Seedlings grew only slowly during winter, and most severely photoinhibited plants grew the slowest. Correlations between growth and photoinhibition as measured during winter persisted into spring even though pre-dawn Fv/Fm made a substantial recovery in all plants.
Spring growth of Eucalyptus polyanthemos appears to be adversely affected by cold-induced photoinhibition over winter. A loss in photosynthetic effectiveness reduces carbon gain during spring, and recovery presumably extends over many weeks, hence a persistence of slower growth despite recovery in Fv/Fm.
In practical terms, establishment of eucalypt seedlings in frost-prone areas benefits from a reduction in irradiance over winter. By analogy, cold-induced photoinhibition could also be responsible for patterns of seedling establishment under parent trees as noted originally.
References
Ball MC, Hodges VC, Laughlin GP (1991) Cold-induced photoinhibition limits regeneration of snow gums at tree line. Funct Ecol 5: 663–668
Holly C, Laughlin GP, Ball MC (1994) Cold-induced photoinhibition and design of shelters for establishment of eucalypts in pasture..Aust J Bot 42: 139–147.