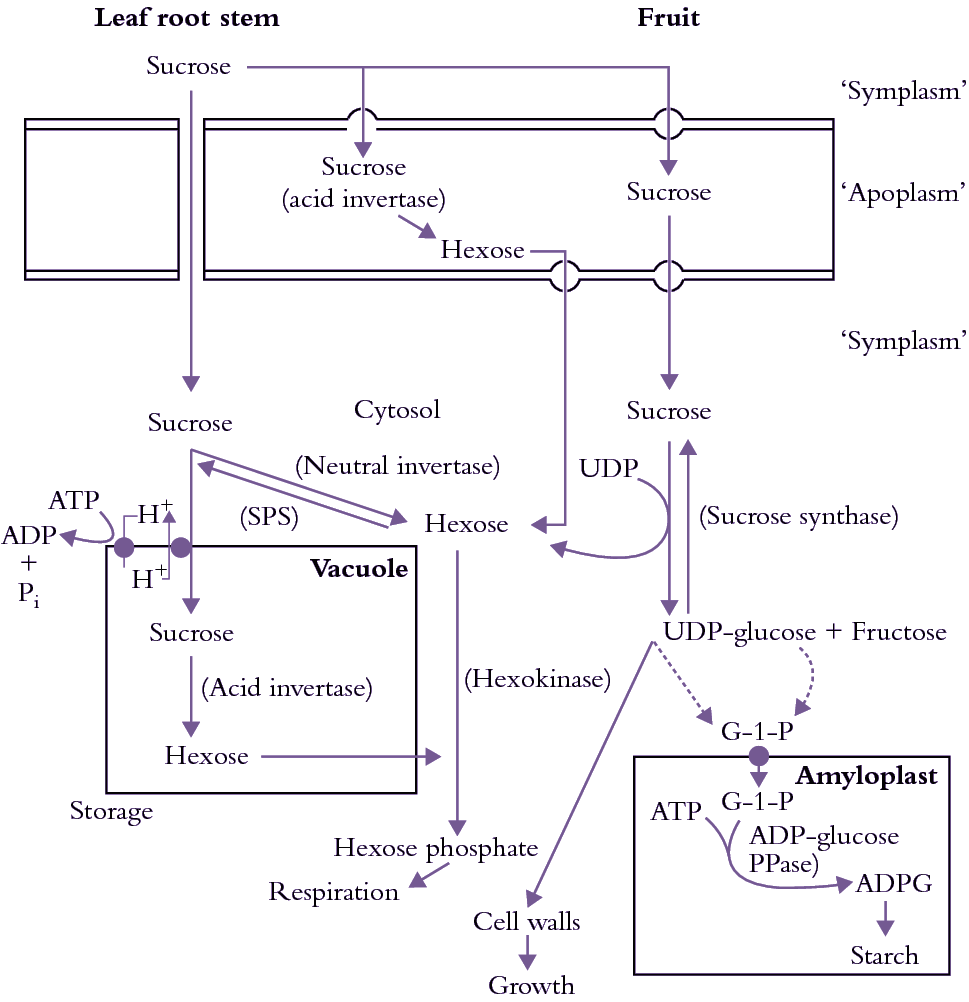
Phloem-imported sucrose can reach the cytoplasm of recipient sink cells chemically unaltered or be hydrolysed en route by extracellular invertase into its hexose moieties. These sugars may then enter a number of metabolic pathways or be compartmented to vacuolar storage (Figure 5.25).
(a) Sucrose metabolism
Sucrose is metabolically inert and, in order to be metabolised, must be hydrolysed to glucose and fructose. Only two enzymes are capable of metabolising sucrose in green plants. These are invertase and sucrose synthase (Figure 5.25) and they are paramount in sugar metabolism after phloem unloading.
Invertase catalyses irreversible hydrolysis of sucrose to its hexose moieties, glucose and fructose. Both acid and neutral invertases occur in plants, with pH optima of about 5 and 7.5, respectively. The activity of invertases varies with plant species, organ type and stage of development. Acid invertases, located in cell wall or in vacuole, are usually active in rapidly growing leaves, stems and fruits and seeds (Ruan et al. 2010), making hexoses available for regulating gene expression and for respiration and biosynthesis. Reduced acid invertase activity in vacuoles during development of sugar cane stems, and its absence from sucrose-accumulating tomato fruit, is a major factor in sucrose accumulation in vacuoles of these tissues. Suppression of cell wall invertase activity led to shrunken seed in maize and small fruit in tomato and loss of pollen fertility in tomato, wheat and rice, demonstrating its critical roles in these reproductive organs. Less is known about the physiological role of neutral invertases..
Sucrose synthase is mainly located in the cytoplasm but recent research also shows that the enzyme may also be associated with plasma membrane and even present in cell wall matrix. It catalyses sucrose cleavage to fructose and UDP-glucose, a high-energy ester of glucose. UDP-glucose is a substrate for biosynthesis of cellulose and may be converted further for starch synthesis.High activities of sucrose synthase are found in both growing and starch storage tissues. In the cytoplasm of starchy tissues, UDP-glucose is converted by UDP-glucose pyro-phosphorylase to glucose-1-phosphate, which is transported across amyloplast membranes. In amyloplasts, glucose-1-phosphate provides glucose moieties for starch synthesis in a pathway comparable to starch formation in chloroplasts of photosynthetic leaves. The critical role of sucrose synthase in starch synthesis is demonstrated with potatoes transformed with an antisense construct of the gene encoding tuber-specific sucrose synthase. Tuber sucrose synthase activity in transformed plants was depressed significantly while the activities of key starch biosynthetic enzymes were unaltered. Low sucrose synthase activity was directly responsible for a proportional decrease in starch accumulation (Zrenner et al. 1995).
Sinks that accumulate soluble sugars have predictably low sucrose synthase activities. Contrastingly high sucrose synthase activities in phloem vessels may be responsible for energy production for phloem loading or unloading and maintaining cellular function of companion cells.
(b) Hexose metabolism
Hexoses transported to the sink cytoplasm are rapidly phosphorylated to hexose-6-phosphates by glucose- and fructose kinases. In these forms, hexoses can be used as substrates for respiration or for synthesis of new cell constituents. Alternatively, sucrose phosphate synthetase can convert them to sucrose, as in leaves (Chapters 1 and 2). Sucrose synthesized by this reaction can be accumulated in vacuoles (e.g. sugar beet tap roots, sugar cane stems) or be rehydrolysed into hexoses by a vacuolar acid invertase (e.g. grape berries).