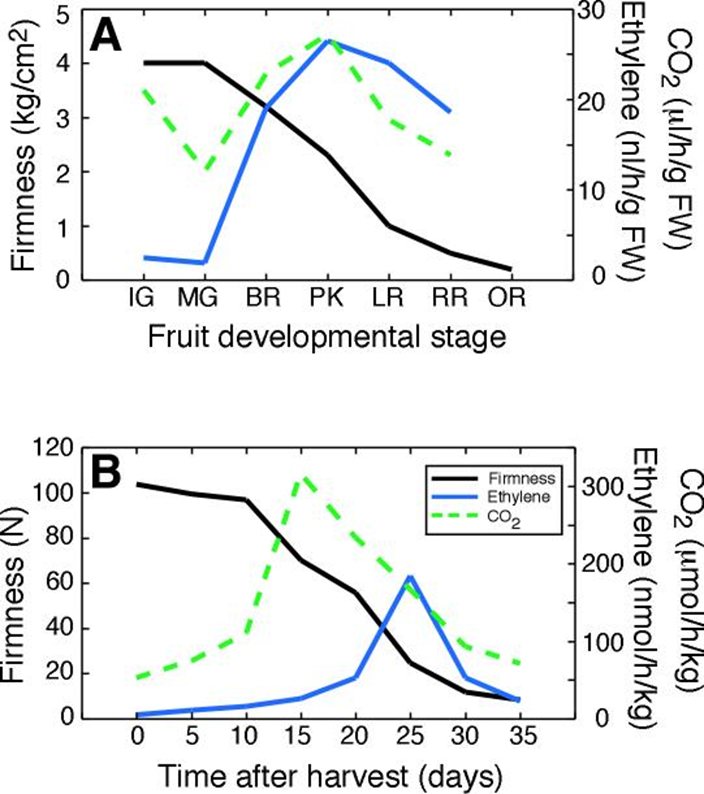
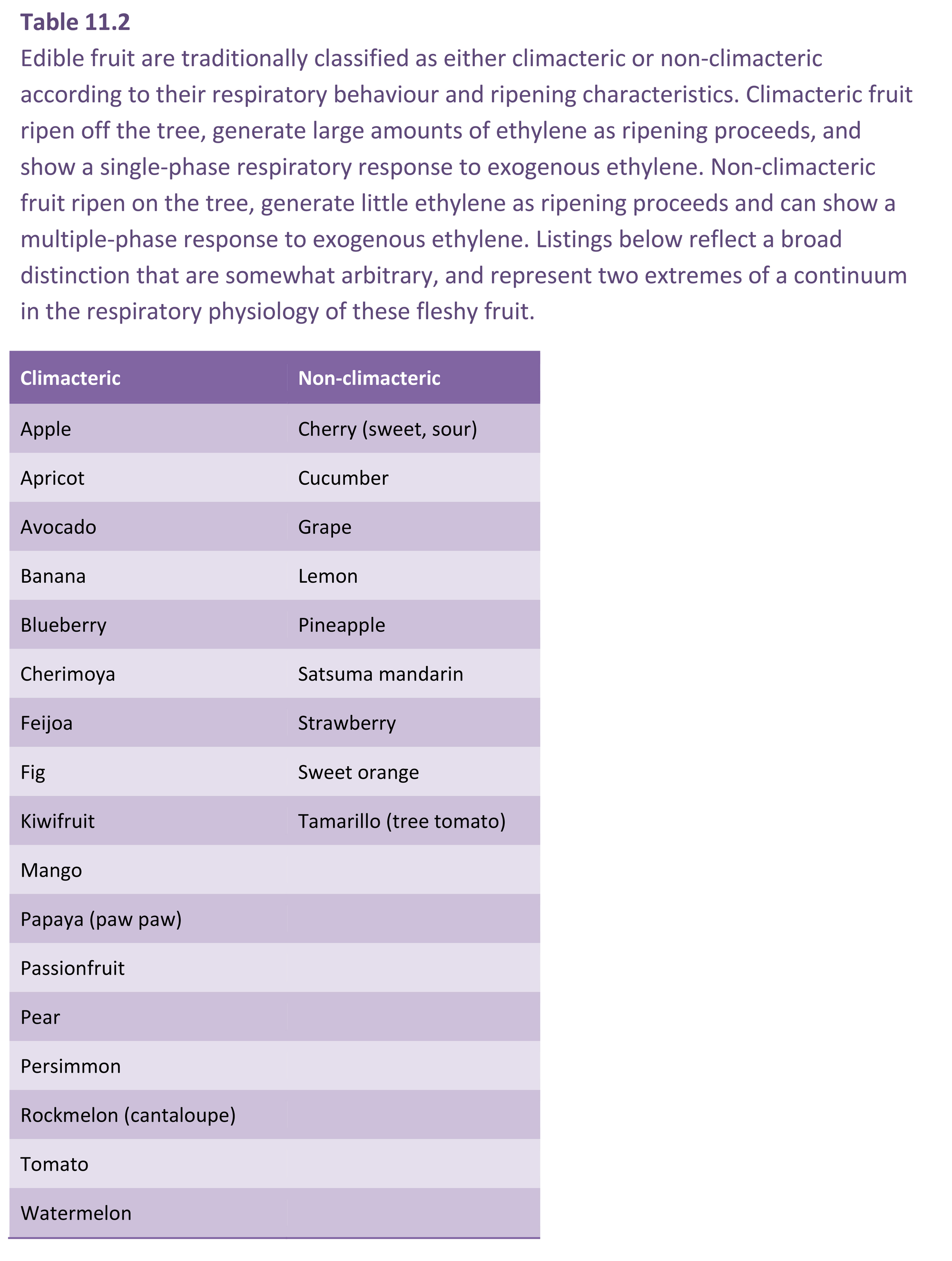
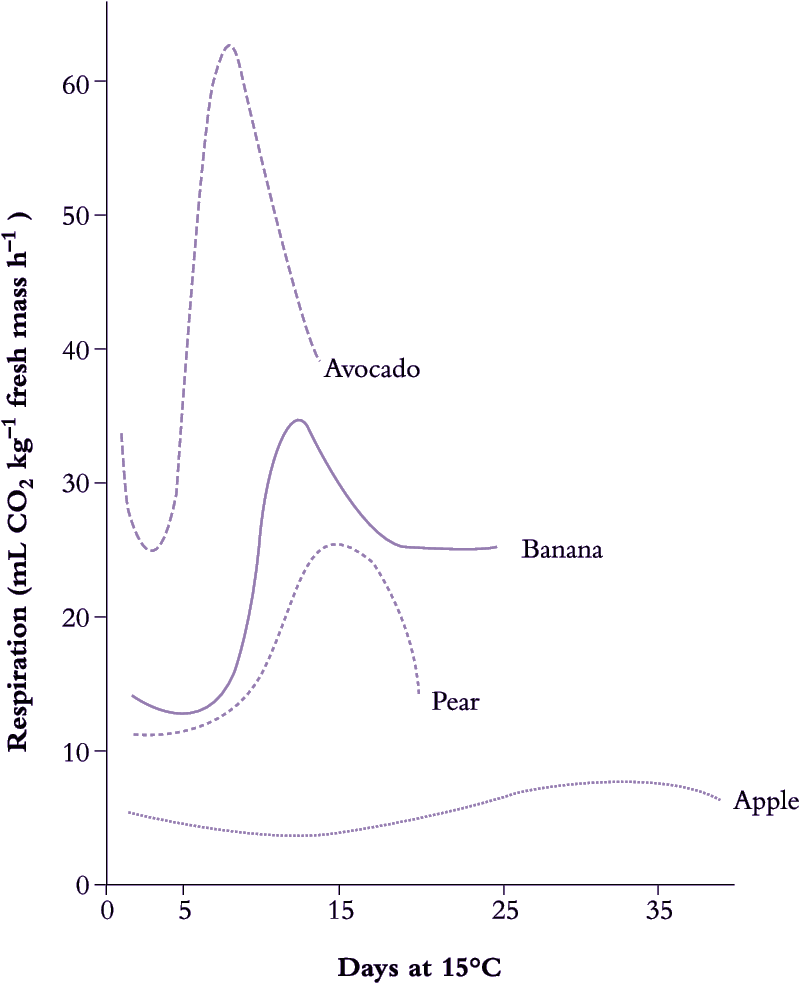
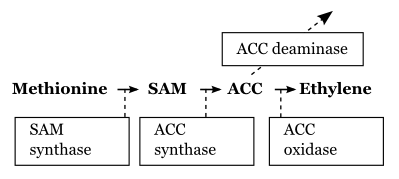
Further reading
Alexander L, Grierson D (2002) Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening. J Exp Bot 53: 2039-2055 http://jxb.oxfordjournals.org/content/53/377/2039.full
Barry CS, Giovannoni JJ (2007) Ethylene and fruit ripening. J Plant Growth Regul 26: 143-159
Brummell DA (2006) Cell wall disassembly in ripening fruit. Funct Plant Biol 33: 103-119
Brummell DA, Harpster MH (2001) Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol Biol 47: 311-340
Gillaspy G, Ben-David H, Gruissem (1993) Fruits: a developmental perspective. Plant Cell 5: 1439-1451 http://www.plantcell.org/content/5/10/1439.full.pdf+html
Kader AA (2008) Flavor quality of fruits and vegetables. J Sci Food Agric 88: 1863-1868
Klee HJ, Giovannoni JJ (2011) Genetics and control of tomato fruit ripening and quality attributes. Annu Rev Genet 45: 41-59
Lee SK, Kader AA (2000) Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol Technol 20: 207-220
Matas AJ, Gapper NE, Chung MY et al. (2009) Biology and genetic engineering of fruit maturation for enhanced quality and shelf-life. Curr Opin Biotechnol 20: 197-203
Paul V, Pandey R, Srivastava GC (2012) The fading distinctions between classical patterns of ripening in climacteric and non-climacteric fruit and the ubiquity of ethylene - An overview. J Food Sci Technol 49: 1-21
Seymour GB, Østergard L, Chapman NH et al. (2013) Fruit development and ripening. Annu Rev Plant Biol 64: 219-241
Literature cited
Atkinson RG, Sutherland P, Johnston SL et al. (2012) Down-regulation of POLYGALACTURONASE1 alters firmness, tensile strength and water loss in apple (Malus x domestica) fruit. BMC Plant Biol 12: 129 (doi:10.1186/1471-2229-12-129) . http://www.biomedcentral.com/1471-2229/12/129
Barry CS, Llop-Tous MI, Grierson D (2000) The regulation of 1-aminocyclopropane-1-carboxylic acid synthase gene expression during the transition from system-1 to system-2 ethylene synthesis in tomato. Plant Physiol 123, 979-986 http://www.plantphysiol.org/content/123/3/979.full
Brummell DA, Harpster MH, Civello PM, Palys JM, Bennett AB, Dunsmuir P (1999) Modification of expansin protein abundance in tomato fruit alters softening and cell wall polymer metabolism during ripening. Plant Cell 11: 2203-2216 http://www.plantcell.org/content/11/11/2203.full
Dahmani-Mardas F, Troadec C, Boualem A, Leveque S, Alsadon AA, Aldoss AA, Dogimont C, Bendahmane A (2010) Engineering melon plants with improved fruit shelf life using the TILLING approach. PLoS ONE 5: e15776. doi: 10.1371/journal.pone.0015776
Dai N, Cohen S, Portnoy V et al. (2011) Metabolism of soluble sugars in developing melon fruit: a global transcriptional view of the metabolic transition to sucrose accumulation. Plant Mol Biol 76: 1-18
Davies C, Wolf T, Robinson SP (1999) Three putative sucrose transporters are differentially expressed in grapevine tissue. Plant Sci 147: 93-100
de Jong M, Mariani C, Vriezen WH (2009) The role of auxin and gibberellin in tomato fruit set. J Exp Bot 60: 1523-1532 http://jxb.oxfordjournals.org/content/60/5/1523.full
Espley RV, Brendolise C, Chagné D et al. (2009) Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. Plant Cell 21: 168-183 http://www.plantcell.org/content/21/1/168.full
Ferreira SA, Pitz KY, Manshardt R et al. (2002) Virus coat protein transgenic papaya provides practical control of Papaya ringspot virus in Hawaii. Plant Disease 86: 101-105
Hackett RM, Ho CW, Lin Z et al. (2000) Antisense inhibition of the Nr gene restores normal ripening to the tomato Never-ripe mutant, consistent with the ethylene receptor-inhibition model. Plant Physiol 124: 1079-1085 http://www.plantphysiol.org/content/124/3/1079.full
Harada S, Fukuta S, Tanaka H, Ishiguro Y, Sato T (1995) Genetic analysis of the trait of sucrose accumulation in tomato fruit using molecular marker. Breed Sci 45: 429-434
Harker FR, Redgwell RJ, Hallett IC et al. (1997) Texture of fresh fruit. Hortic Rev 20: 121-224
Johnston JW, Gunaseelan K, Pidakala P et al. (2009) Co-ordination of early and late ripening events in apples is regulated through differential sensitivities to ethylene. J Exp Bot 60: 2689-2699 http://jxb.oxfordjournals.org/content/60/9/2689.full
Kevany BM, Tieman DM, Taylor MG et al. (2007) Ethylene receptor degradation controls the timing of ripening in tomato fruit. Plant J 51: 458-467
Klann EM, Hall B, Bennett AB (1996). Antisense acid invertase (TIV1) gene alters soluble sugar composition and size in transgenic fruit. Plant Physiol 112: 1321-1330 http://www.plantphysiol.org/content/112/3/1321.abstract
Kobayashi S, Goto-Yamamoto N, Hirochika H (2004) Retrotransposon-induced mutations in grape skin color. Science 304: 982.
Lichtenthaler HK, Rohmer M, Schwender J (1997) Two independent biochemical pathways for isopentenyl diphosphate and isoprenoid biosynthesis in higher plants. Physiol Plant 101: 643-652
McAtee P, Karim S, Schaffer R, David K (2013) A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front Plant Sci 4: 79 http://www.frontiersin.org/Plant_Cell_Biology/10.3389/fpls.2013.00079/abstract
Mworia EG, Yoshikawa T, Salikon N et al. (2012) Low-temperature-modulated fruit ripening is independent of ethylene in 'Sanuki Gold' kiwifruit. J Exp Bot, 63: 963-971 http://jxb.oxfordjournals.org/content/63/2/963.full
Quesada MA, Blanco-Portales R, Posé S et al. (2009) Antisense down-regulation of the FaPG1 gene reveals an unexpected central role for polygalacturonase in strawberry fruit softening. Plant Physiol 150: 1022-1032 http://www.plantphysiol.org/content/150/2/1022.full
Saladié M, Matas AJ, Isaacson T et al. (2007) A re-evaluation of the key factors that influence tomato fruit softening and integrity. Plant Physiol 144: 1012-1028 http://www.plantphysiol.org/content/144/2/1012.full
Sevillano L, Sanchez-Ballesta MT, Romojaro F, Flores FB (2009) Physiological, hormonal and molecular mechanisms regulating chilling injury in horticultural species. Postharvest technologies applied to reduce its impact. J Sci Food Agric 89: 555-573
Shackel KA, Greve C, Labavitch JM, Ahmadi H (1991) Cell turgor changes associated with ripening in tomato pericarp tissue. Plant Physiol 97: 814-816 http://www.plantphysiol.org/content/97/2/814.full.pdf+html
Sisler EC, Serek M (1997) Inhibitors of ethylene responses in plants at the receptor level: Recent developments. Physiol Plant 100: 577-582
Sitrit Y, Bennett AB (1998) Regulation of tomato fruit polygalacturonase mRNA accumulation by ethylene: a re-examination. Plant Physiol 116: 1145-1150 http://www.plantphysiol.org/content/116/3/1145.full
Souleyre EJF, Greenwood DR, Friel EN et al. (2005) An alcohol acyl transferase from apple (cv. Royal Gala), MpAAT1, produces esters involved in apple fruit flavor. FEBS J 272: 3132-3144
Stitt M, Sonnewald U (1995) Regulation of metabolism in transgenic plants. Annu Rev Plant Physiol Plant Mol Biol 46: 341-368
Tacken E, Ireland H, Gunaseelan K et al. (2010) The role of ethylene and cold temperature in the regulation of the apple POLYGALACTURONASE1 gene and fruit softening. Plant Physiol 153: 294-305 http://www.plantphysiol.org/content/153/1/294.full
Tanaka Y, Sasaki N, Ohmiya A (2008) Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids. Plant J 54: 733-749
Walker AR, Lee E, Bogs J et al. (2007) White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J 49: 772-785
Watkins CB (2008) Overview of 1-methylcyclopropene trials and uses for edible horticultural crops. Hort Sci 43: 86-94
Yokotani N, Nakano R, Imanishi S et al. (2009) Ripening-associated ethylene biosynthesis in tomato fruit is autocatalytically and developmentally regulated. J Exp Bot 60: 3433-3442 http://jxb.oxfordjournals.org/content/60/12/3433.full
Zhang XY, Wang XL, Wang XF et al. (2006) A shift of phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. Plant Physiol 142: 220-232 http://www.plantphysiol.org/content/142/1/220.full