11.3.2 - Composition of photoassimilates
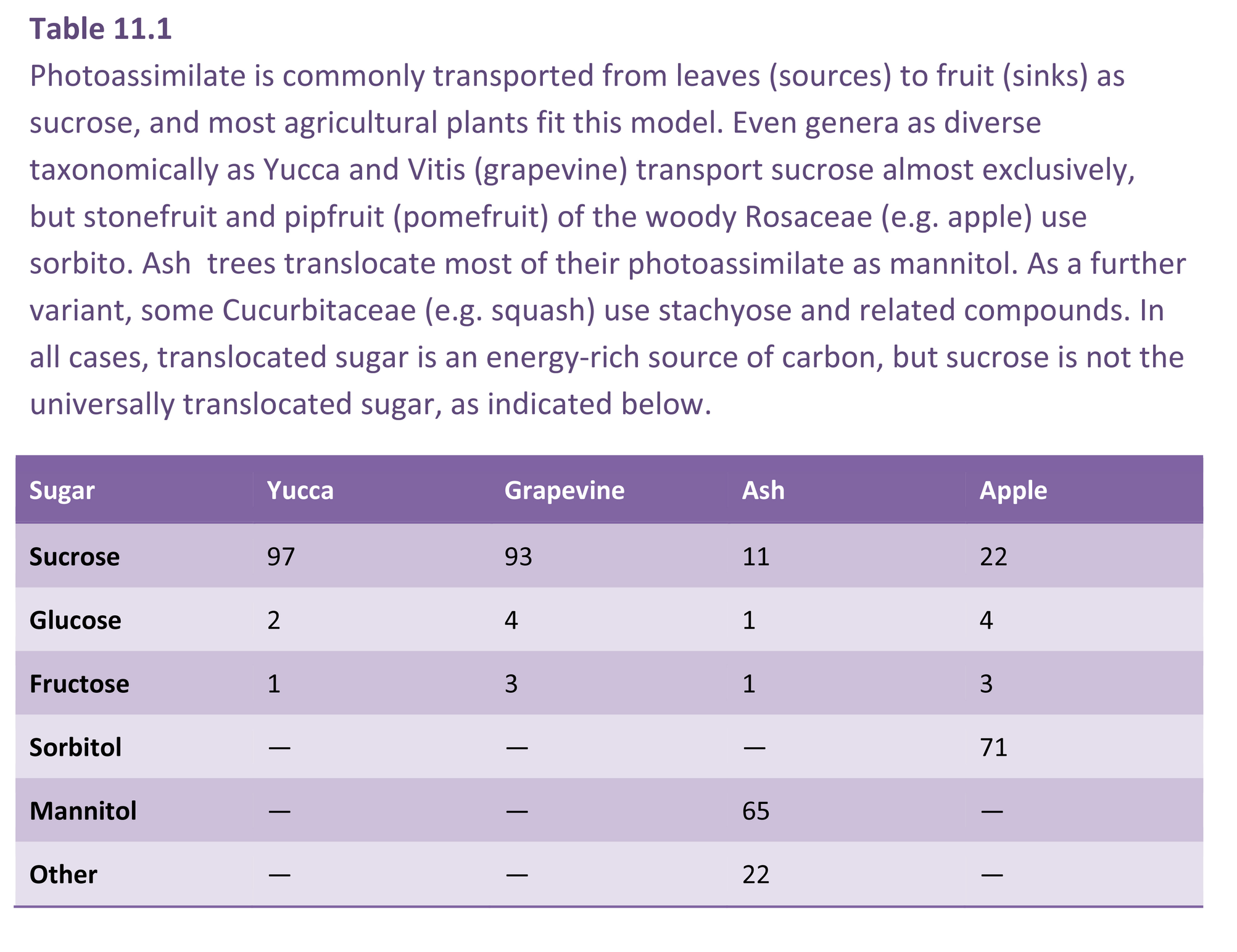
Radiolabelling of photoassimilates has also been used to identify which compounds are transported into storage organs. Analyses of phloem tissues and phloem sap show that in most plants carbohydrate enters fruits primarily as sucrose. However, other soluble carbohydrates can predominate in some plants of commercial importance (Table 11.1).
In the woody Rosaceae (apple, pear, stonefruit), the sugar alcohol sorbitol is the major photosynthetic product at 60–85% of transported carbon, the remainder being mainly sucrose. Regardless of transport form, photoassimilate arriving in fruits is rapidly converted to the storage products characteristic of the fruit in question (principally starch, glucose, fructose and sucrose). Thus the identity of labelled sugars in fruit often differs markedly from the form transported. For example, sorbitol concentration is high during early development of apple fruit and more or less reflects the composition of photoassimilate in transit. By maturity, sorbitol content will typically decline to below 5% of the total soluble carbohydrate.
If sorbitol reaching fruit is not fully metabolised, apoplastic accumulation results and pericarp tissues become glassy in a disorder called ‘watercore’ (see below; Figure 11.23). This is a common problem with some apple cultivars such as Fuji. Sugar transport and accumulation can thus have economic importance — both in terms of desired taste characteristics and postharvest fruit quality.
In kiwifruit, the polyol myo-inositol may comprise up to 35% of soluble carbohydrate in developing fruit, and up to 20% in leaves. As yet, we do not know whether inositol, like sorbitol, is transported in the phloem, or whether there may be physiological disorders caused by inadequate metabolism of sorbitol within fruit. Such findings challenge our common perception of sucrose as the universal transport carbohydrate in economic crops, and suggest that we still have a lot to learn about the control of carbohydrate metabolism.