During fruit ripening, softening and textural changes (including the development of juiciness) are components of the suite of modifications that make ripe fruit attractive to animals that might disperse the seeds. The texture of ripe fruit differs drastically between species, with crisp, hard apple and deformably soft avocado representing the extremes. The characteristic textures of different fruit and their manner of softening can be linked both with anatomical features and with changes that occur to the cell wall during ripening (Figure 11.15). Some fruit that are picked while hard, such as kiwifruit and tomato, will subsequently soften markedly as a result of extensive modifications to the cell wall structure that include substantial swelling. Other fruit, such as apple or watermelon, remain crisp and soften only slightly. Their thin cell walls remain relatively unaltered. Both types of softening occur in the pear family: Asian pear (Nashi) shows a crisp apple-like texture, whereas many European pears soften to give ripe fruit with a melting texture. Interspecific crosses between the two types show that texture is heritable (Harker et al. 1997).
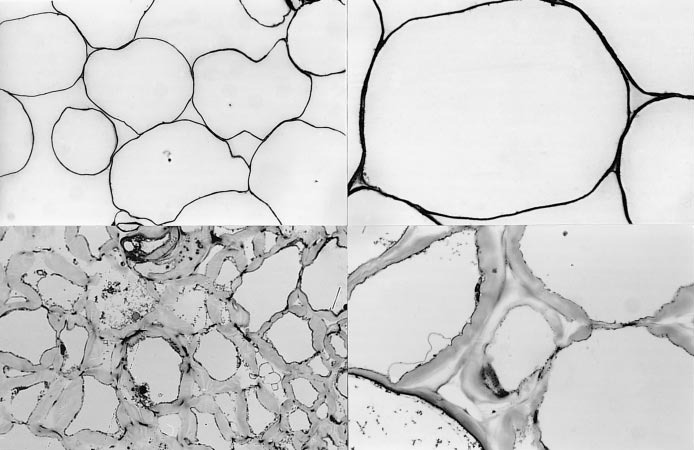
Many textural characteristics relate to the fate of fruit flesh when it is fractured and crushed in the mouth. Contributing factors include cell size, cell adhesion, turgor and packing, wall thickness, wall composition and the reaction of cells to shearing stress as they are chewed (Harker et al. 1997). For example, a ripe apple has large (0.1–0.3 mm diameter), turgid, thin-walled cells that are loosely packed (airspace c. 20% of fruit volume). When that flesh is chewed, cells fracture and release their sugary contents as free juice. In contrast, ripe kiwifruit has minimal airspace (c. 2% of fruit volume) and cell walls are thick and hydrophilic (Figure 11.15). Such cells tend to pull apart when the flesh is chewed, resulting in a paste moistened by liquid held in cell walls or released by damaged cells. Avocado also has cells with walls that are thick and soft and which tend to pull apart, but also has a high proportion of oil that gives the pulp an oily quality in the mouth.
Softening in kiwifruit occurs over a period of weeks, and can be divided into a number of phases (Figure 11.16). Modification of the cell wall plays an important part in determining fruit texture and ripening characteristics. The plant primary cell wall consists of a network of strong, rigid cellulose microfibrils held together by a complex matrix of polysaccharides consisting of two types: the hemicelluloses (composed mainly of neutral sugars) and the pectins (rich in galacturonic acid), together with smaller amounts of structural proteins. The outer part of each cell wall, which abuts and provides attachment to neighbouring cells, is composed mainly of pectins and is called the middle lamella. Fruit softening involves alterations to various pectin and hemicellulose polysaccharide wall components, and changes to the bonding between some polymers. Wall modification has been the subject of much research worldwide, mostly using tomato as a model, but also using other fruit in the search for common themes (Brummell 2006). Chemical analyses of cell wall components in a range of species, notably kiwifruit and tomato, show some consistent changes during the early stages of ripening. In kiwifruit, these include:
- Solubilisation of pectin (but without further degradation);
- The cell wall swells and shows an increased affinity for water (become more hydrophilic);
- Loss of galactose from pectins (especially of a galactan that is tightly associated with the cellulose microfibrils);
- De-esterification of some pectins.
These changes continue once kiwifruit have begun rapidly softening to ripeness (phase 2 in Figure 11.16). Phase 2 softening is associated with a further increase in pectin solubilisation, loss of galactan and arabinan side chains from pectic polymers, and more cell wall swelling. As softening progresses into phase 3 two more important changes begin, both of which appear to be regulated by ethylene:
- Depolymerisation (a reduction in size) of the hemicellulosic polysaccharide xyloglucan, which is associated with a reduction in cell wall strength;
- Depolymerisation of pectin, which is associated with dissolution of the middle lamella and reduced intercellular adhesion.
These six changes have been observed in a wide range of fruit types, although the extent and relative timing varies somewhat between species. Such observations indicate that pectin solubilisation and cell wall swelling are important events in the control of softening in kiwifruit and probably most other species with a melting texture. Cell wall modification is much less extensive in fruit with a crisp, fracturable texture such as apple and capsicum pepper. As fruit become fully ripe, dissolution of the middle lamella means that it eventually virtually disappears as a visible structure under the microscope. Dissolution of the middle lamella results in a great reduction in intercellular adhesion, and cells now have fewer regions of attachment to each other and become more rounded in appearance as they pull away from neighbouring cells. The primary walls are also weakened by the various changes that have occurred, and cells easily rupture when bitten or chewed, releasing the cell contents as juice.
Many of the cell wall-modifying enzymes produced during ripening are regulated by ethylene. Polygalacturonase (an enzyme that depolymerises pectins) increases de novo 10–50-fold in tomato fruit during ripening (Sitrit and Bennett 1998). Understandably, this enzyme was originally accorded a major role in the ripening process. However, studies with transgenic tomato fruit in which polygalacturonase was suppressed found only a small reduction in softening during ripening, although there was a very substantial increase in the storage life of the fruit (Figure 11.17). Because transgenic fruits retained firmness for longer, they were left on vines longer, resulting in more carbohydrate accumulation prior to harvest. Moreover, fruit could be harvested partially coloured rather than mature green, thereby allowing ripening processes to progress more naturally and yielding fruit with better flavour and appearance. To move from experimental results to public availability, the fruit had to go through a series of tests and be de-regulated. Following USDA approval, a transgenic cultivar producing fruit with >99% reduction in polygalacturonase activity was named Flavr SavrTM by Calgene, and was released for marketing in the USA under the brand identity of McGregor. Tomato paste with increased viscosity derived from similar transgenic fruit was successfully marketed in the UK for several years.
The effects of reduced polygalacturonase activity on firmness and shelf life were probably largely due to decreased degradation of the middle lamellae, and thus the maintenance of intercellular connections and fruit integrity.
Although this work was originally interpreted as suggesting a very minor role for polygalacturonase in fruit softening, the relatively small reduction in firmness observed may have been due to two factors. Firstly, tomato has atypically high levels of polygalacturonase enzyme (far more than in other species), and secondly silencing of the polygalacturonase gene was incomplete, meaning that in this species even 0.5% of wild-type activity was still substantial. Subsequently, silencing of polygalacturonase was found to partially but significantly reduce fruit softening in strawberry and apple, thus confirming that pectin depolymerisation is one part of the softening process (Quesada et al. 2009; Atkinson et al. 2012). What these studies have also clearly demonstrated is that softening is not controlled by a single cell wall-modifying enzyme. Rather, many different enzymes are involved, with enzymes such as polygalacturonase, pectate lyase, expansin, β-galactosidase and pectin methylesterase making specific contributions to the softening process (Brummell and Harpster 2001; Brummell 2006). It is the action of these many enzymes working together that brings about the wall swelling, reduced wall strength and weakened middle lamellae that result in the final softening and textural characteristics of the ripe fruit. Indeed, the actions of the various enzymes may be interdependent. For example, polygalacturonase requires the prior de-methylesterification of pectin by pectin methylesterase to make the substrate susceptible (Brummell and Harpster 2001), and the action of expansin to increase the accessibility of enzyme to substrate in the cell wall (Brummell et al. 1999).
In addition to cell wall disassembly, work in several species has shown that a decrease in cellular turgor accompanies fruit ripening and is an important component of softening (Shackel et al. 1991). This is caused partly by internal water movements resulting from the movement of solutes from symplast to apoplast, and partly to the loss of water from the fruit. In tomato, analysis of the non-softening DFD mutant attributed its enhanced postharvest firmness to very low water loss from the fruit and therefore to cellular turgor being maintained at higher levels than in wild-type (Saladié et al. 2007).