Temperature
The earliest attempts at temperature management were dependent on fruit being held in cold caves, or using cold night air to prolong the storage life, but experience showed that a ‘best’ temperature can be sharply defined, and may differ between species or even cultivars (Sevillano et al. 2009). To obtain the maximum benefit from cold temperatures, the temperature must be as low as possible without causing damage to the fruit; this is termed the lowest safe temperature. Below the lowest safe temperature, but at non-freezing temperatures, the fruit may develop symptoms of chilling injury (See Section 14.4). At even lower temperatures, generally in the range –0.5°C to –1.5°C, freezing occurs which irreversibly damages a living product. Because of this, –0.5°C is usually the lowest temperature used for storage of fruit, including some apple cultivars, berries or 'Hayward' kiwifruit. Temperatures at which chilling symptoms occur are around 8oC for subtropical species and may be anything up to 14°C for some tropical fruit: for example unripe banana and mango need to be shipped at 13–14°C. However, it is not only tropical and sub-tropical fruit that are susceptible to chilling injury; even 'Hayward' kiwifruit, which is stored at 0°C or just below, may develop chilling injury.
At 0°C, respiration is reduced to a level that is just enough to maintain cell function. Sugar is slowly consumed during this process so that fruit with a low sugar content at harvest are less durable. Commodities such as kiwifruit, which are picked with large supplies of carbohydrate in the form of starch, have an additional source of sugar to utilise, giving longer storage lives than those entirely reliant on soluble reserves, such as grapes.
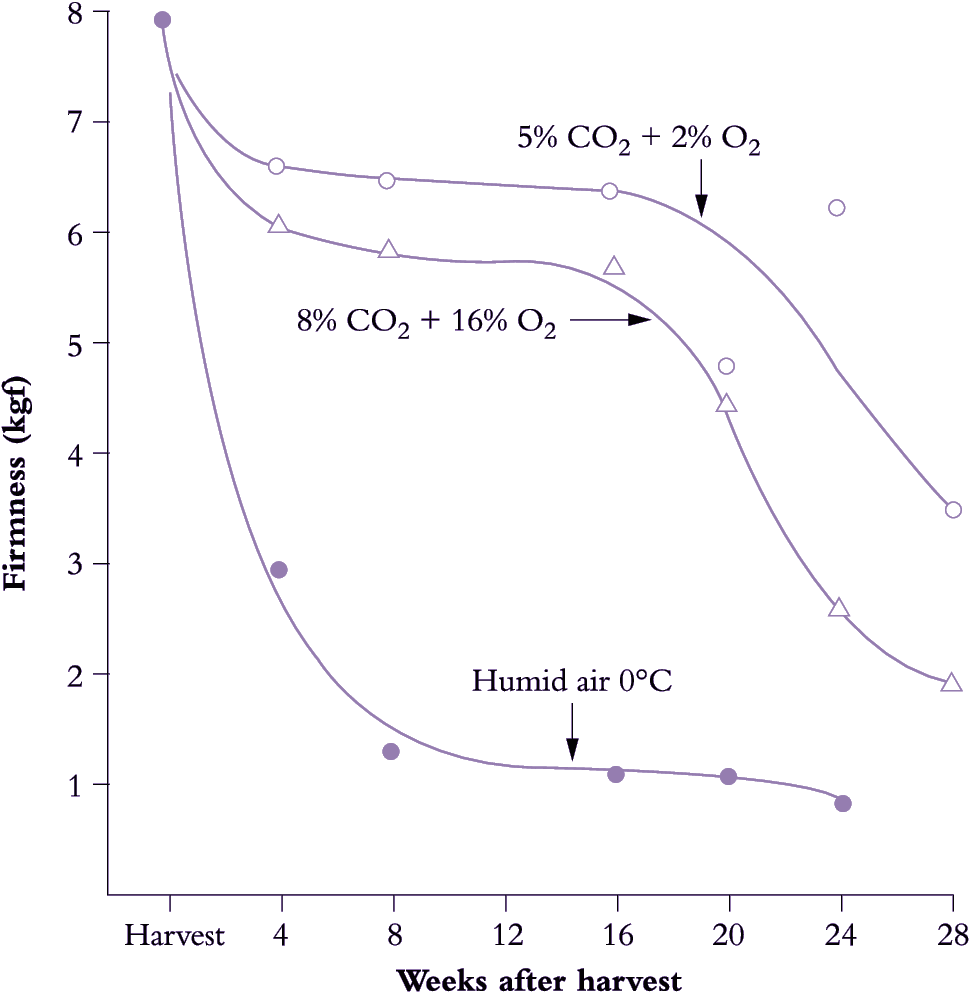
Low-temperature storage has played an important part in the development of successful fruit export industries in Australasia, because of the great shipping distances between orchard and consumer. The success of kiwifruit has been largely due to its ability to be stored at 0°C for 6 months or more with no detrimental effect on flavour or texture.
Associated with low-temperature storage is a wide range of techniques to manage temperature changes en route to storage (Kader 2002). There are strong differences between species in their temperature management requirements. Elements of temperature management that need to be considered include the timing of cooling after harvest, the rate of cooling and the final storage temperature. Temperature management may also be viewed as a two-stage process, the removal of the field heat and then temperature maintenance during storage. While it is generally considered that the field heat should be removed from fruit as soon as possible after harvest, there are circumstances where delays may be advantageous for the postharvest performance of the fruit. So whilst highly perishable berryfruit tend to be cooled as soon as possible after harvest, kiwifruit and some stonefruit benefit from a delay at ambient temperature before cooling. Exactly what happens during this delay period is not clear; it may simply be a continued progress of fruit development or the loss of a small amount of water. However, the delay tends to make the fruit more tolerant of storage at low temperatures. In the case of 'Hayward' kiwifruit, the delay period is termed ‘curing’ and is specifically applied to reduce the incidence of stem-end rots caused by Botrytis. As a beneficial side effect, the low temperature tolerance of the fruit is also increased. In this sense, curing in kiwifruit is not the same as the curing for wound healing of the skin that is commonly referred to for sweet potatoes.
The rate of cooling is dependent both on what is required commercially and what can be tolerated by the fruit. Simply placing fruit, either in bulk bins or packed, in a coolstore will result in the fruit being cooled, the rate of which will depend on the initial fruit temperature, the cooling capacity of the refrigeration equipment, the airflow in the store and any insulating effects from the packaging, especially if the fruit are packed in boxes with polyliners and held on pallets. The rate of cooling can be increased by forced air cooling, also termed precooling, in which cold air is actively drawn past the fruit. This is a rapid method for removal of field heat, after which temperature management in a coolstore removes the smaller heat load that results from continued respiratory activity during storage. In some cases fast precooling may induce high incidences of chilling damage. This is one reason why 'Hayward' kiwifruit is not always precooled, but may be cooled from about 14–18°C at harvest to about 2°C after about 5 days, with a further 5–7 days to reach the final storage temperature.
Managing the rate of cooling of fruit to avoid chilling injury may be as simple as allowing the fruit to cool slowly, as in the case of 'Hayward' kiwifruit described above, or there may be clearly defined stages of cooling whereby fruit are cooled to an intermediate temperature, held for a period of days before the temperature is reduced to the final storage temperature. In all these instances of slow cooling, there is a trade off between the conditioning effect that increases tolerance to low temperatures and the progression of fruit development that occurs more rapidly at higher temperatures, and reduces storage life of the fruit.
An extreme example of temperature treatment prior to storage is where fruit may be treated at high temperatures (40–50°C) for disinfestation, and in particular to kill fruit fly, after which the fruit ripening may be slower than would occur naturally.
The expression of chilling injury symptoms may be reduced in long-term storage by intermittently warming the fruit. However, whilst there are numerous reports of such treatments in the scientific literature, the practicalities of the procedure and detrimental side effects to fruit quality make it commercially uncommon.
Relative humidity
Once harvested, fruit will continuously lose water to a point where quality will be affected. In some species, a small amount of water loss may accelerate ripening (e.g. avocado), but in all fruit there eventually comes a point at which loss of water, usually first seen as shrivelling, results in the fruit becoming unacceptable. Water loss from the fruit is driven by the vapour pressure gradient between the fruit and the surrounding environment. While the capacity for air to hold water is reduced at low temperatures, there is always a gradient driving water from the fruit into the coolstore atmosphere. The less fruit there is in a coolstore, the greater the water loss from each fruit before an equilibrium relative humidity is reached. Water may be lost from the coolstore atmosphere by condensation on the refrigeration coils that are colder than the room atmosphere, and the greater the temperature differential between the coils and atmosphere the greater the loss of water. When storage is at about 0°C, this can be seen by ice developing on the coils that must be removed by defrosting.
In preventing quality loss of harvested fruit, the relative humidity of the storage environment is one of the first aspects considered, since fruit will lose water more rapidly at lower relative humidity. This is mostly an issue where fruit are held unpacked or in bulk in a coolstore, and water loss is exacerbated where there is only a small volume of fruit in the store, air flow is high and there is a large temperature differential on the refrigeration coil. In other circumstances, such as for kiwifruit that may be stored for months, the fruit is packed into fibreboard packs with a polyethylene liner or bag. In these circumstances, it is the bag that creates a high humidity environment for the fruit and limits the fruit’s water loss. A very high relative humidity in the store environment where packed fruit are held may be detrimental to the integrity of the fibreboard packaging, which would soften and lose its strength.