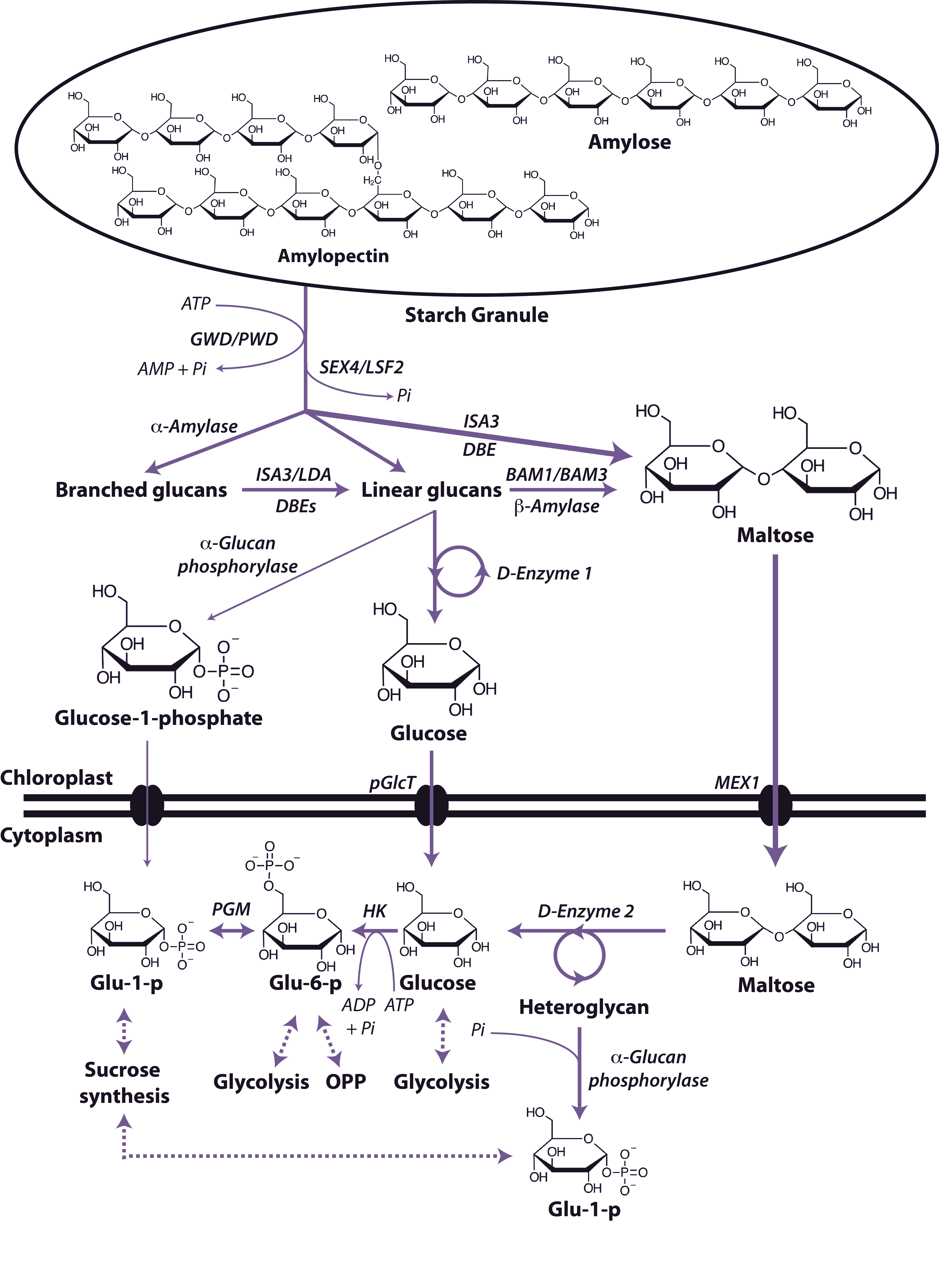
Starch is the principal storage carbohydrate in plants and this carbon reserve plays a number of important roles in plants. It is composed of two polymers of glucose, amylose and amylopectin and is stored in the plastid (chloroplast in leaves, amyloplasts in non-photosynthetic tissues) as insoluble, semi-crystalline granules. Starch is accumulated during rapid growth in the day and is almost completely degraded at night to mostly glucose and maltose, which is exported from the chloroplast and metabolised in the cytosol (Figure 2.19). Starch degradation is initiated by the addition of phosphate groups at the C6-position and C3-position of individual glucosyl residues that act to disrupt the packing of the glucans at the granule surface. These phosphate additions are catalysed by two enzymes, glucan water dikinase (GWD) and phosphoglucan water dikinase (PWD) respectively. The hydrolysis of the resulting glucan and phosphoglucan chains is carried out by a suite of enzymes including the phosphoglucan phosphatases (SEX4/LSF2), β-amylases (BAM1/BAM3), debranching enzymes (DBE; ISA3/LDA), α-amylase (AMY3), α-glucan phosphorylase and the disproportionating enzyme 1 (D-enzyme 1; an α-1,4-glucanotransferase).). The resulting maltose and glucose are exported to the cytosol by the glucose transporter (pGlcT) and maltose transporter (MEX1) and glucose-1-phosphate is thought to be exported by a similar but yet unknown mechanism. Once in the cytosol the maltose and glucose are converted to substrates for either sucrose synthesis, glycolysis or the oxidative pentose phosphate pathway by a number of enzymes including the disproportionating enzyme 2 (D-enzyme 2; an α-1,4-glucanotransferase), α-glucan phosphorylase, hexokinase and phosphoglucomutase.
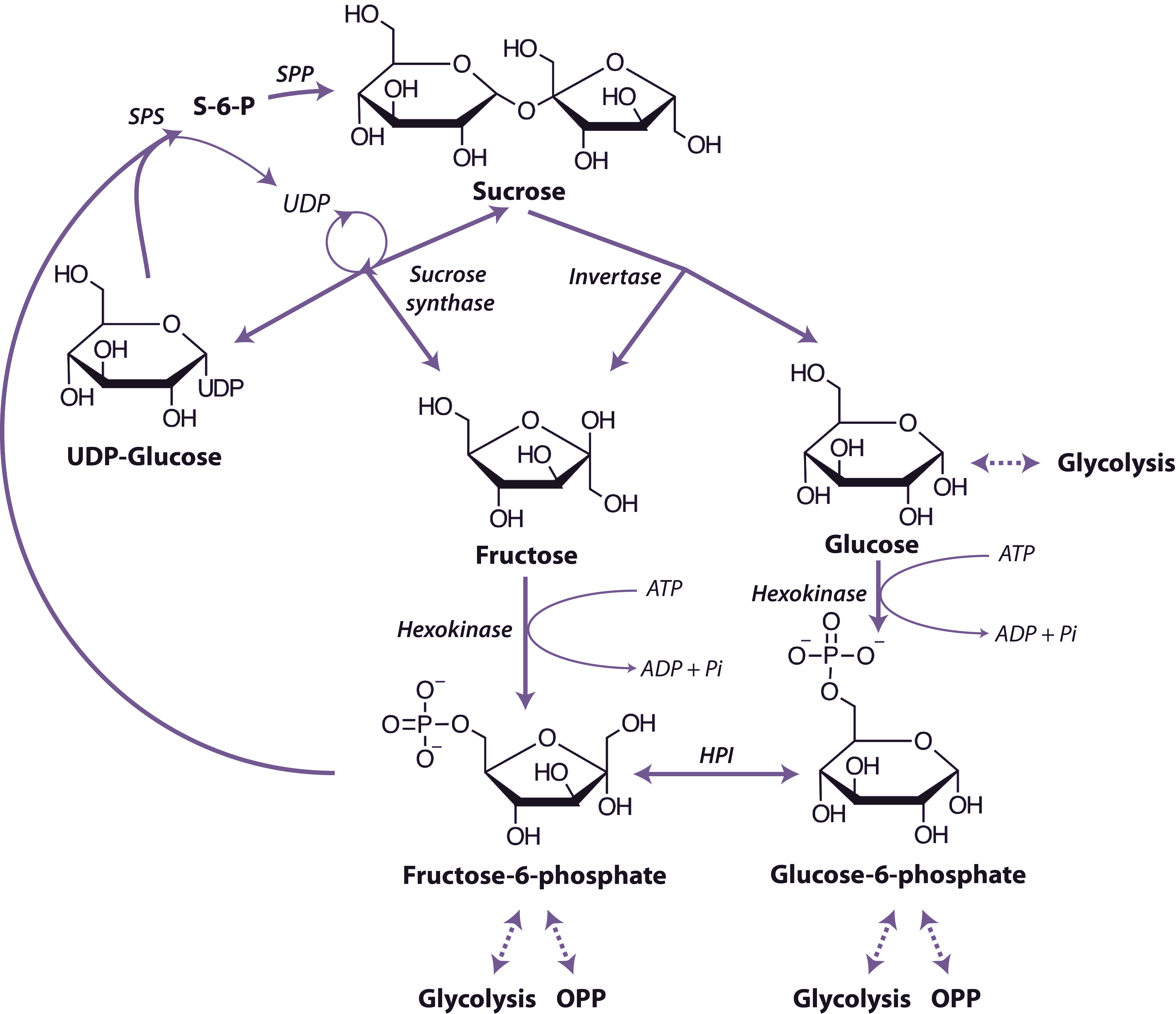
Sucrose is the world’s most abundant disaccharide, it is only produced by photosynthetic organisms and serves a role as a transportable carbohydrate and sometimes as a storage compound. The reactions in plant tissues leading to degradation of sucrose to hexose monophosphates are outlined in Figure 2.20.
The first step is cleavage of the glycosidic bond by either invertase (Equation 2.1) or sucrose synthase (Equation 2.2).
\[\text{Sucrose + H}_2\text{O} \rightarrow \text{D-Glucose + D-Fructose}\tag{2.1}\]
\[\text{Sucrose + UDP} \rightarrow \text{UDP-Glucose + D-Fructose}\tag{2.2}\]
Plant tissues contain distinct invertases located in the vacuole, cell wall (acid invertases) cytosol, mitochondria, nucleus, and cholorplast (neutral/alkaline invertases) which hydrolyse sucrose to glucose and fructose in an irreversible reaction. The invertases are differentially regulated by a number of mechanisms including pH to allow them to function in cell expansion, supply of carbon skeletons and energy metabolism. Multiple isoforms of sucrose synthase are located in the cytosol or cytosolic membranes that catalyse a thermodynamically reversible reaction, but this reaction probably acts only to breakdown sucrose in vivo. Their activity is developmentally regulated and they have functions in the supply of activated glucose for starch and cellulose biosynthesis. While both invertase and sucrose synthase can both breakdown sucrose, research using knockouts of multiple isoforms of both enzymes has shown that sucrose synthase is not required for normal growth in Arabidopsis, whereas invertase is indispensable. However this does not rule out the requirement of sucrose synthase in certain tissues of crop plant tubers, seeds and fruits where it has been shown to be crucial. Glucose and fructose are metabolised further by phosphorylation to the corresponding hexose-6-P by hexokinase. Hexokinase in plant tissues is associated with the outer surface of mitochondria.