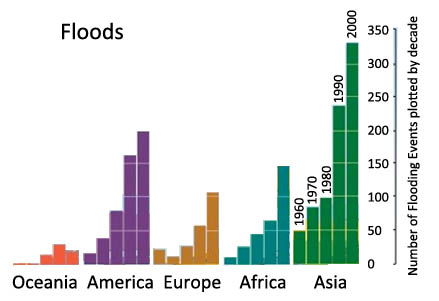
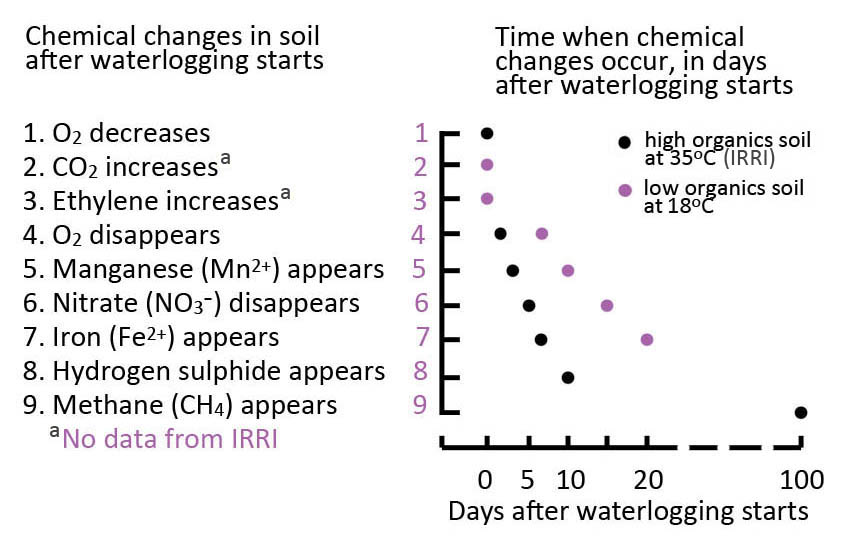
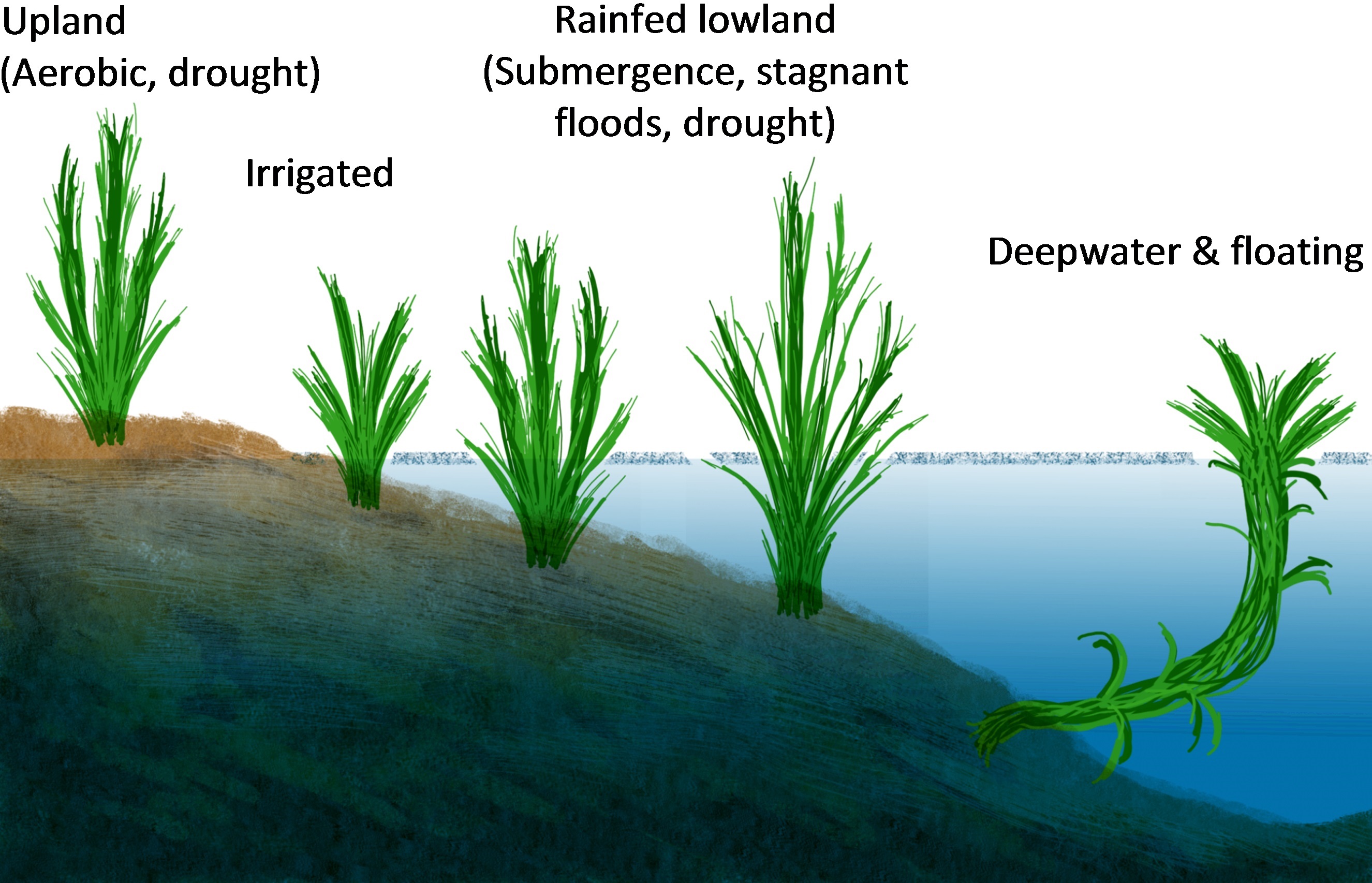
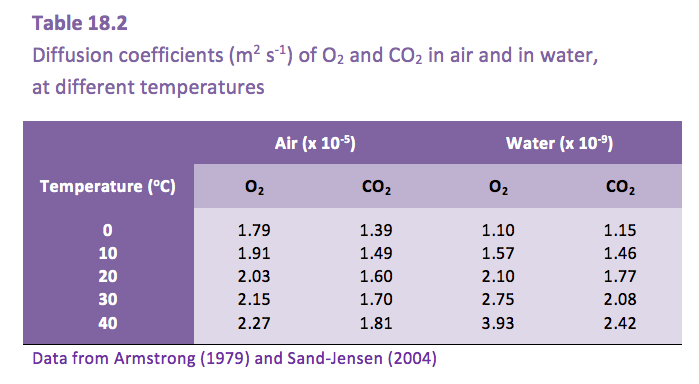
Armstrong W (1979) Aeration in higher plants. Adv Bot Res 7: 225-332
Armstrong W, Armstrong J, Beckett PM, Justin SHFW (1991) Convective gas-flows in wetland plant aeration. In: Jackson MB, Davies DD, Lambers H (eds). Plant life under oxygen deprivation. The Hague: SPB Academic Publishing. pp. 283-302.
Bailey-Serres J, Voesenek LACJ (2008) Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 59: 313–339
Beckett PM, Armstrong W, Justin SHFW, Armstrong J (1998) On the relative importance of convective and diffusive gas-flows in plant aeration. New Phytol 110: 463-468
Blokhina O, Virolainen E, Fagerstedt KV (2003) Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot 91: 179-194
Brix H, Sorrell BK, Orr PT (1992) Internal pressurization and convective gas flow in some emergent freshwater macrophytes. Limnol Oceanogr 37: 1420-1433
Carystinos GD, Macdonald HR, Monroy AF et al. (1995) Vacuolar H+-translocating pyrophosphatase is induced by anoxia or chilling in seedlings of rice. Plant Physiol 108: 641-649
Chen Z, Cuin TA, Zhou M et al. (2007) Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. J Exp Bot 58: 4245-4255
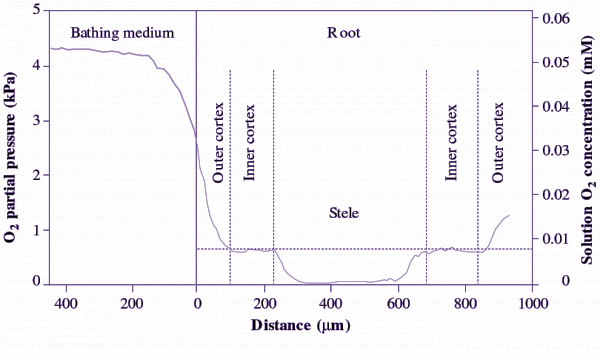
Colmer TD (2003) Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ 26: 17-36
Colmer TD, Greenway H (2011) Ion transport in seminal and adventitious roots of cereals during O2 deficiency. J Exp Bot 62: 39-57
Dacey JWH (1980) Internal winds in water lilies: an adaptation for life in anaerobic sediments. Science 210: 1017-1019
Das KK, Sarkar RK, Ismail AM (2005) Elongation ability and non-structural carbohydrate levels in relation to submergence tolerance in rice. Plant Sci 168: 131-136
Demidchik V, Cuin TA, Svistunenko D et al. (2010) Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: single-channel properties, genetic basis and involvement in stress-induced cell death. J Cell Sci 123: 1468-1479
Demidchik V, Shabala SN, Davies JM (2007) Spatial variation in H2O2 response of Arabidopsis thaliana root epidermal Ca2+ flux and plasma membrane Ca2+ channels. Plant J 49: 377-386
Drew MC, Saglio PH, Pradet A (1985) Larger adenylate energy charge and ATP/ADP ratios in aerenchymatous roots of Zea mays in anaerobic media as a consequence of improved internal oxygen transport. Plant Physiol 165: 51-58
Ella ES, Kawano N, Ito O (2003a) Importance of active-scavenging system in the recovery of rice seedlings after submergence. Plant Sci 165: 85-93
Ella E, Kawano N, Yamauchi Y et al. (2003b) Blocking ethylene perception enhances flooding tolerance in rice. Funct Plant Biol 30: 813-819
Elzenga JTM, van Veen H (2010) Waterlogging and plant nutrient uptake. In: Mancuso S, Shabala S (eds) Waterlogging signalling and tolerance in plants Springer-Verlag, Heidelberg, pp 23-36
Felle HH (2010) pH signaling during anoxia. In: Mancuso S, Shabala S (eds) Waterlogging signalling and tolerance in plants. Springer-Verlag, Heidelberg, pp 79-98
Gibbs J, Greenway H (2003) Mechanism of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Funct Plant Biol 30: 1-47
Greenway H, Gibbs J (2003) Mechanisms of anoxia tolerance in plants. II. Energy requirements for maintenance and for energy consuming processes. Funct Plant Biol 30: 999-1036
Himmelblau DM, Arends E (1959) Die Löslichkeit inerter Gase in Wasser bei hohen Temperaturen und Drucken. Chemie Ingenieur Technik 31: 791-795
Ismail AM, Heuer S, Thomson M, Wissuwa M (2007) Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol Biol 65:547–570
Jackson MB, Armstrong W (1999) Formation of aerenchyma and the processes of plant ventilation in relation to soil flooding and submergence. Plant Biol 1: 274-287
Jackson MB (2003) The Impact of Flooding Stress on Plants and Crops. PlantStress.Com. Ed. A Blum http://www.plantstress.com/articles/waterlogging_i/waterlog_i.htm
Johnson JR, Cobb BG, Drew MC (1989) Hypoxic induction of anoxia tolerance in roots of Zea mays. Plant Physiol 91: 837-841
Johnson JR, Cobb BG, Drew MC (1994) Hypoxic induction of anoxia tolerance in roots of Adh1 null Zea mays L. Plant Physiol 105: 61-67
Justin SHFW, Armstrong W (1987) The anatomical characteristics of roots and plant response to soil flooding. New Phytol 106: 465-495
Khabaz-Saberi H, Rengel Z, Wilson R, Setter TL (2010) Variation of tolerance to manganese toxicity in Australian hexaploid wheat. J Plant Nutr Soil Sci 173: 103-112
Lee SC, Mustroph A, Sasidharan R et al. (2011) Molecular characterization of the submergence response of the Arabidopsis thaliana ecotype Columbia. New Phytol 190: 457-471.
Mackill DJ, Ismail AM, Singh US et al. (2012) Development and rapid adoption of submergence-tolerant (Sub1) rice varieties. Adv Agron 115: 303-356
Malik AI, Colmer TD, Lambers H et al. (2002) Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol 153: 225-236
Morrell S, Greenway H, Davies DD (1989) Regulation of pyruvate decarboxylase in vitro and in vivo. J Exp Bot 41: 131-139
Noctor G, Foyer CH (1998) Ascorbate and glutathione: Keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49: 249-279
Pang J, Shabala S (2010) Membrane transporters and waterlogging tolerance. In: Waterlogging signalling and tolerance in plants. Mancuso S, Shabala S (eds). Springer-Verlag, Heidelberg, pp 197-219
Pang JY, Cuin T, Shabala L et al. (2007) Effect of secondary metabolites associated with anaerobic soil conditions on ion fluxes and electrophysiology in barley roots. Plant Physiol 145: 266-276
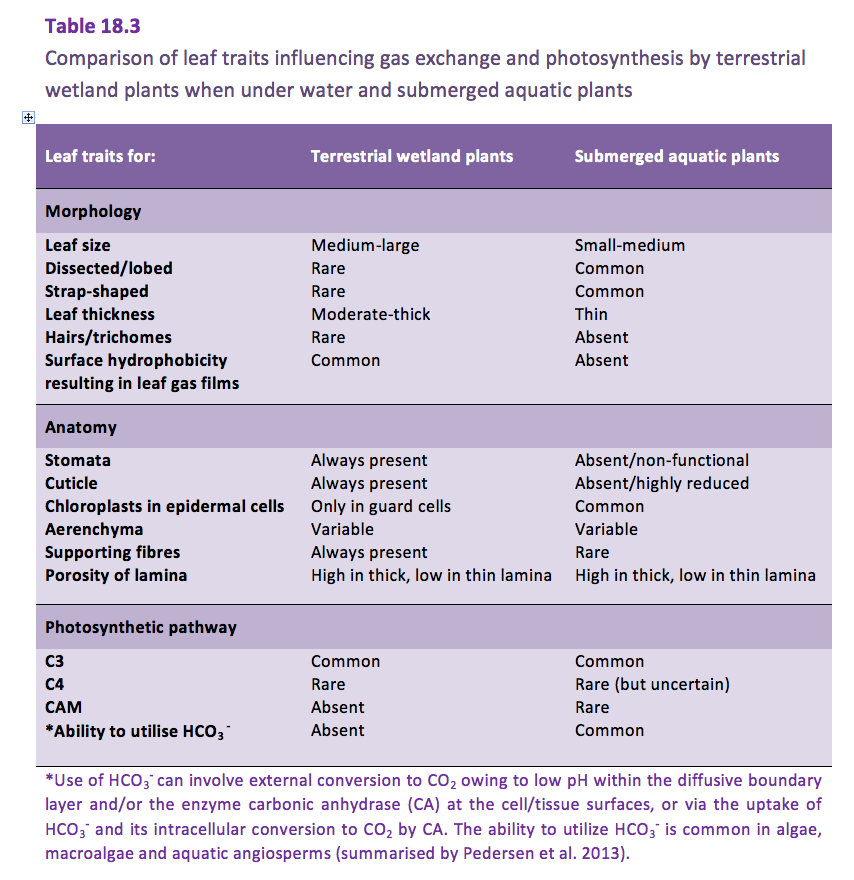
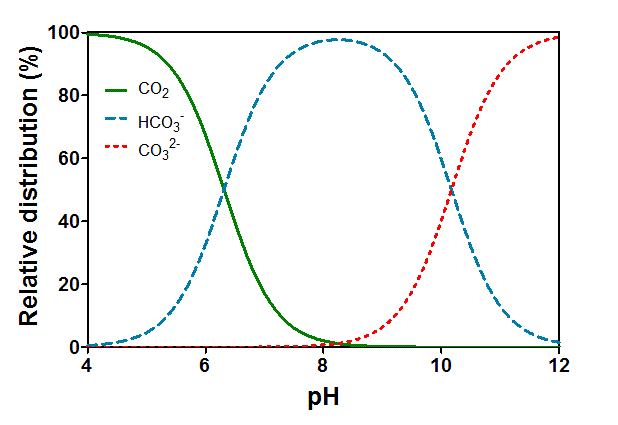
Pedersen O, Colmer TD, Sand-Jensen K (2013) Underwater photosynthesis of submerged plants – recent advances and methods. Front Plant Sci 4 DOI: 10.3389/fpls.2013.0014
Raymond P, Pradet A (1980) Stabilization of adenine nucleotide ratios at various levels by an oxygen limitation of respiration in germinating lettuce seeds. Biochem J 190: 39-44
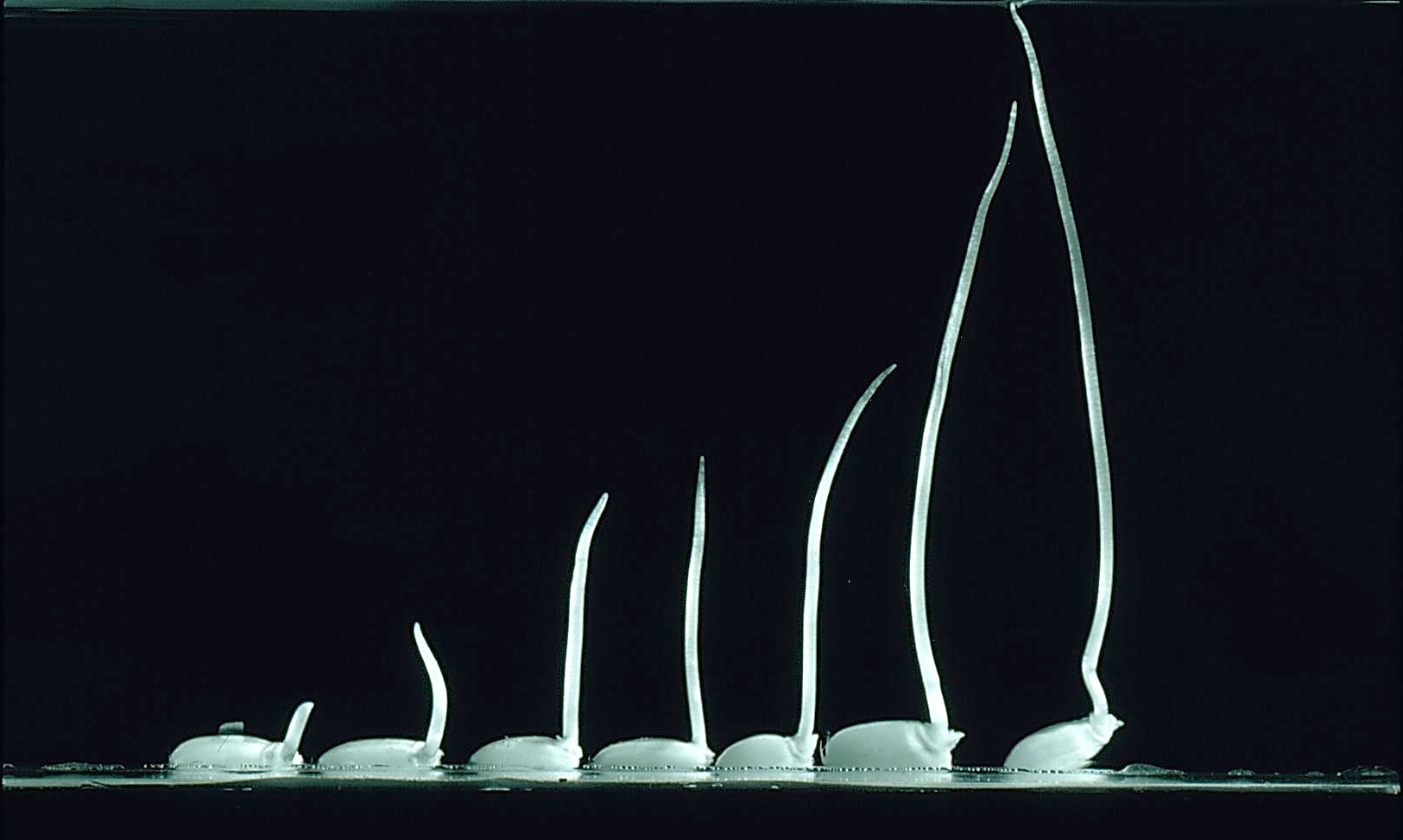
Rich SM, Ludwig M, Colmer TD (2012) Aquatic adventitious root development in partially and completely submerged wetland plants Cotula coronopifolia and Meionectes brownii. Ann Bot 110: 405-414
Sand-Jensen K (2004) Økologi og biodiversitet. Gyldendal, Copenhagen
Schmitz AJ, Folsom JJ, Jikamaru Y et al. (2013) SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway. New Phytol 198:1060-1070
Setter TL, Laureles EV (1996) The beneficial effect of reduced elongation growth on submergence tolerance of rice. J Exp Bot 47: 1551-1559
Shabala S (2003) Regulation of potassium transport in leaves: from molecular to tissue level. Ann Bot 92: 627-634
Shabala S (2011) Physiological and cellular aspects of phytotoxicity tolerance in plants: the role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol 190: 289-298
Sorrell BK, Hawes I (2010) Convective gas flow and the maximum depths achieved by helophyte vegetation in lakes. Ann Bot 105: 165-174
Steffens B, Sauter M (2009) Epidermal cell death in rice is confined to cells with a distinct molecular identity and is mediated by ethylene and H2O2 through an autoamplified signal pathway. Plant Cell 21: 184-196
Stumm W, Morgan JJ (1996) Aquatic chemistry. 3rs ed. New York: John Wiley & Sons.
Thomson CJ, Greenway H (1991) Metabolic evidence for stellar anoxia in maize roots exposed to low O2 concentrations. Plant Physiol 96: 1294-1301
Vashist D, Hesselink A, Pierik R et al. (2011) Natural variation of submergence tolerance among Arabidopsis thaliana accessions. New Phytol 190: 299-310
Visser EJW, Bogemann GM, Blom CWPM, Voesenek LACJ (1996) Ethylene accumulation in waterlogged Rumex plants promotes formation of adventitious roots. J Exp Bot 47: 403-410
Vretare Strand V (2002) The influence of ventilation systems on water depth penetration of emergent macrophytes. Freshwater Biol 47: 1097-1105
Waters I, Morrell S, Greenway H, Colmer TD (1991) Effects of anoxia on wheat seedlings. II. Influence of O2 supply prior to anoxia on tolerance to anoxia, alcoholic fermentation, and sugar levels. J Exp Bot 42: 1437-1447
Wilhelm E, Battino R, Wilcock RJ (1977) Low-pressure solubility of gases in liquid water. Chem Rev 77: 219-262
Yan B, Dai Q, Liu X, Huang S, Wang Z (1996) Flooding-induced membrane damage, lipid oxidation and activated oxygen generation in corn leaves. Plant Soil 179: 261-268
Zhang Q, Greenway H (1994) Anoxia tolerance and anaerobic catabolism of aged beetroot storage tissues. J Exp Bot 45: 567-575