
Early development of a pineapple. Phloem conduits from the leaves distribute sugars to the growing inflorescence, with flower buds arranged in spirals, which will later develop into the large juicy fruit.
Early development of a pineapple. Phloem conduits from the leaves distribute sugars to the growing inflorescence, with flower buds arranged in spirals, which will later develop into the large juicy fruit.
Contributing Authors: Craig Atkins2; Yong-Ling Ruan1. 1University of Newcastle, Australia. 2School of Plant Biology, University of Western Australia
This Chapter is updated from a previous version written by John W Patrick, Ian Wardlaw and Tina Offler for Plants in Action 1st Edition.
A plant is a coordinated network of assimilatory regions (sources) linked to regions of resource utilisation (sinks). The phloem vascular system provides a path for assimilate transport from source to sink.
The phloem conduits distribute the sugars made in the leaves to growing tissues and organs that cannot carry out photosynthesis. These ‘sinks’ include shoot and root apices, flower buds, and developing fruit and seed.
Xylem conduits are responsible for delivery of water, inorganic nutrients and organic forms of nitrogen to transpiring leaves (Chapters 3 and 4).
Section 5.1 describes the pathway of the distribution of sugars made in chloroplasts, as well as nitrogen assimilates made in the leaves, to growing organs and other non-photosynthetic tissues. Section 5.2 describes the composition of phloem sap and how to collect it. Quantitative information is presented on the speed of phloem transport from sources to sinks, and the controls of long-distance transport.
Cellular and regulatory mechanisms of phloem loading in leaves are shown in Section 5.3, and mechanisms of phloem unloading at sinks in Section 5.4 with particular reference to developing seeds.
The focus of this chapter is on the transport of sugars. The transport of amino acids and other nitrogen-containing compounds is equally important, and the same general principles apply to nitrogen-containing or phosphorus-containing compounds that are synthesised in the leaf.
CO2 fixed by photosynthesis in chloroplasts has several possible fates, but most ends up as sucrose or starch. Starch is stored in chloroplasts, and sucrose is stored in vacuoles of mesophyll cells. Both starch and vacuolar sucrose serve as temporary storage pools from which the cytoplasmic sucrose pool is replenished. Sucrose, along with amino acids and mineral nutrients, is loaded into the phloem tissue which consists of sieve element—companion cell (se—cc) complexes for long-distance transport to growing tissues and other non-photosynthetic sinks. These solutes are exchanged reversibly between se-cc complexes and short- and long-term storage pools along the axial pathway. Short-term storage pools include phloem apoplasm, and the protoplasm of non-transport cells provides a long-term storage pool. At the end of the pathway, sucrose and other transported solutes are consumed in respiration and growth, or are stored as solutes in vacuoles or polymers in amyloplasts (starch) or protein bodies.
The overall flow of photoassimilates throughout the plant can therefore be called a source–path–sink system (Figure 5.1).
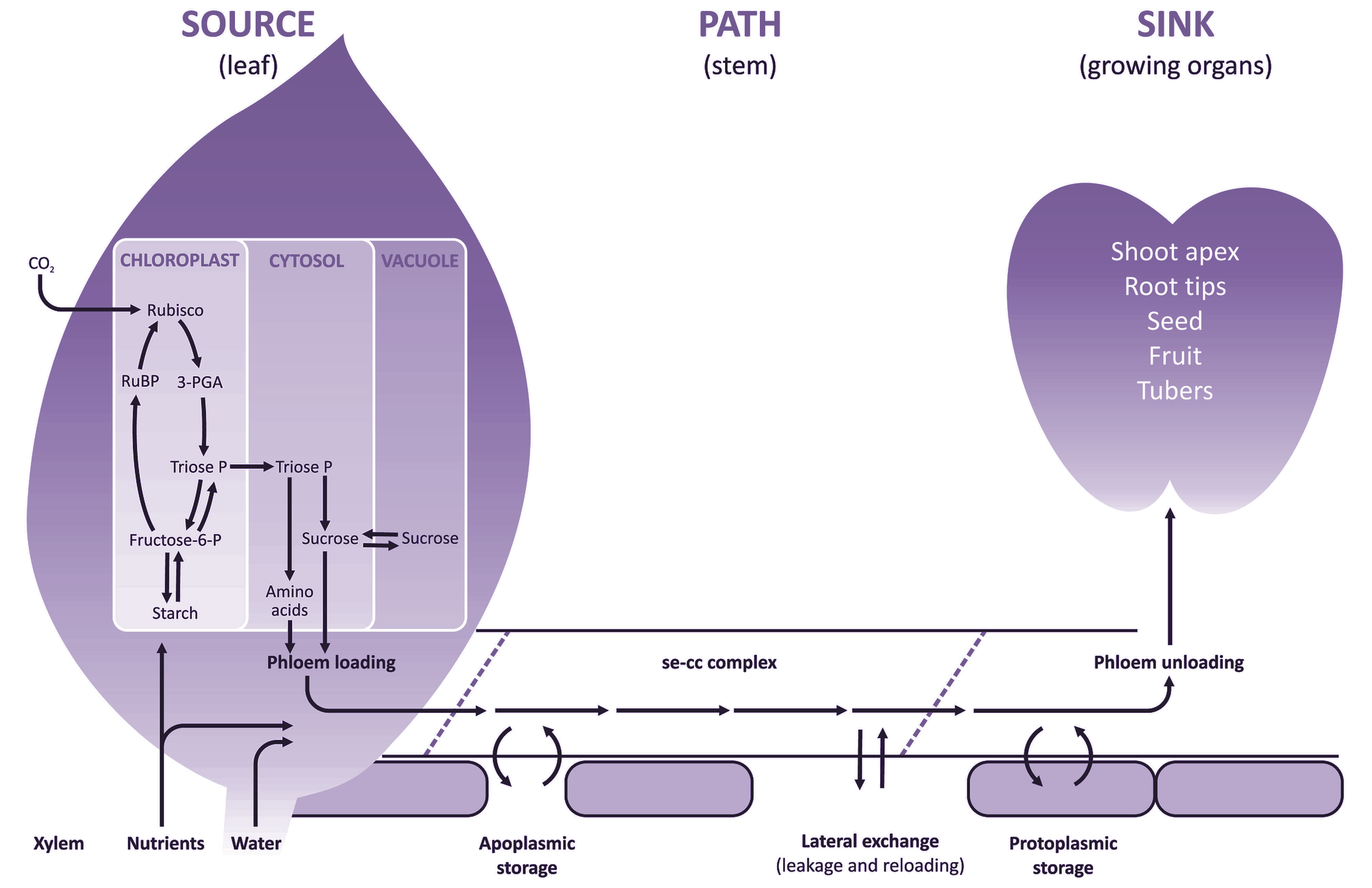
Figure 5.1. Schematic diagram of transfer and transport processes contributing to the flow of assimilates acquired from aerial or soil environments, through the source-path-sink system. CO2 fixed by photosynthesis in chloroplasts gives rise to sucrose and starch. Sucrose, amino acids and mineral nutrients are loaded into sieve element—companion cell (se—cc) complexes of leaf phloem for long-distance transport to non-photosynthetic sinks. These solutes are exchanged reversibly between se-cc complexes and short- and long-term storage pools along the axial pathway. Short-term storage pools include phloem apoplasm, whereas the protoplasm of non-transport cells provides a long-term storage pool. In sink tissues, solutes are used for respiration, growth or storage.
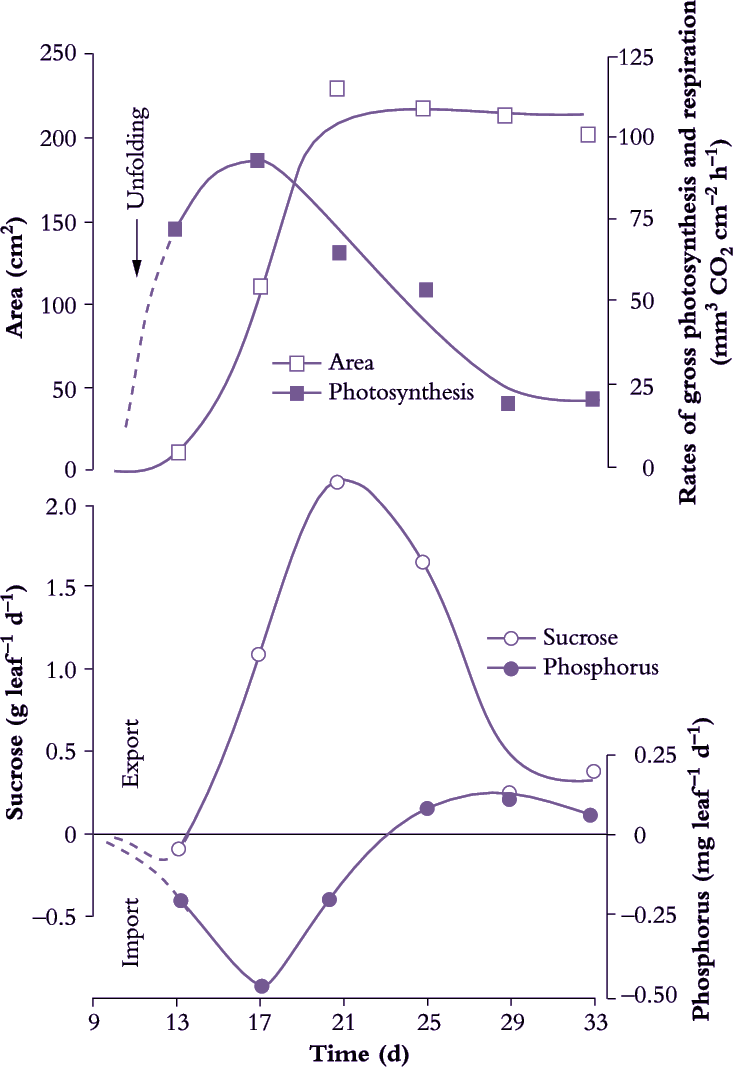
Figure 5.2 Time course of sucrose and phosphorus (P) net import and export from a leaf during its development. As a cucumber leaf expands, net sucrose export coincides with the rise in net leaf photosynthetic rate (O) to meet photoassimilate demands of young leaves. Once a leaf has reached some 30% of its final area, net photosynthesis by the whole leaf exceeds photoassimilate demand by growth and so excess sucrose can be exported. Thereafter, the rate of sucrose export closely follows photosynthetic rate, reaching a maximum when the leaf reaches its final size and gradually declining thereafter. Import of P (and other mineral nutrients) continues throughout leaf expansion and P export only starts once the leaf is fully expanded. Sucrose import and export were calculated from the difference between rates of whole-leaf photosynthesis and dry matter gain (Based on Hopkinson 1964; reproduced with permission of Journal of Experimental Botany)
Net export of photoassimilates occurs from fully expanded leaves (Figure 5.2) and long-term storage pools located along the axial transport pathway. Chloroplasts of C3 plants (Chapters 1 and 2) partition photoassimilates between the photosynthetic oxidative cycle and starch biosynthesis or release them immediately to the cytosol as triose phosphate for sucrose synthesis. In non-starch-forming leaves, high concentrations of sugars can be accumulated in the vacuoles of mesophyll cells or made available for immediate loading into the phloem and export. Leaves also serve as secondary sources for nutrients and amino acids previously delivered in the transpiration stream. Nutrients and amino acids can be exported in the phloem immediately, or after accumulation in short-term storage pools.
An additional source of photoassimilates is located along the axial phloem path (petioles, stems, peduncles, pedicels and roots) as a result of leakage from the vascular tissues. Leaked photoassimilates accumulate in short- or long-term storage pools which serve as secondary sources to buffer photo-assimilate supplies to the sinks against shifts in export rates from the primary photoassimilate sources.
Assimilates including sucrose, amino acids are transferred into sieve elements of fully expanded leaves against significant concentration and electrochemical gradients. This process is referred to as phloem loading. The cellular pathways of phloem loading, and hence transport mechanisms and controls, vary between plant species. Longitudinal transport of assimilates through sieve elements is achieved by mass flow and is termed phloem translocation. Mass flow is driven by a pressure gradient generated osmotically at either end of the phloem pathway, with a high concentration of solutes at the source end and a lower concentration at the sink end. At the sink, assimilates exit the sieve elements and move into recipient sink cells where they are used for growth or storage. Movement from sieve elements to recipient sink cells is called phloem unloading. The cellular pathway of phloem unloading, and hence transport mechanisms and controls, vary depending upon sink function.
Many sink organs are characterised by low rates of transpiration (an exception is a developing leaf) so that most assimilates are delivered by the phloem. Having reached the sink cell cytoplasm through the post-sieve-element transport pathway, assimilates are either metabolised to satisfy the energy, maintenance and growth requirements of sink cells or are compartmented into polymer or vacuolar storage. Collectively, metabolism and compartmentation create a demand for assimilates which is ultimately responsible for driving phloem import.
Sink and source strength must be in balance at a whole-plant level. Thus, an increase in whole-plant sink strength must be matched by an equal increase in source strength, either through increases in source activity or source size. Prior to canopy closure in a crop, much of the increase in source strength comes from increased source size, source activity remaining relatively constant. Significantly, until a leaf has reached some 30% of its final size, photoassimilates for leaf production are exclusively imported through the phloem from fully expanded leaves (Figure 5.2).
During domestication of crop plants, plant breeders selected for crop yield via maximum investment into harvested organs (mostly seeds). Total plant biomass production of advanced wheat is the same as its wild progenitors yet grain yield has increased some 30-fold through breeding. That is, whole-plant source and sink strength have not changed. Increases in wheat yield are associated with a diversion of photoassimilates from vegetative organs to the developing grain, as illustrated by the relative accumulation of 14C photoassimilates exported from the flag leaf.
Final grain yield is not only determined by partitioning of current photoassimilates, but also depends upon remobilisation of non-structural carbohydrates stored in stems, particularly under conditions where environmental stress impairs leaf photosynthesis (Wardlaw 1990). In fact, remobilisation of reserves affects yield in many food plants. For example, deciduous fruit trees depend entirely on remobilised photo-assimilates to support flowering and fruit set as do early stages of pasture regrowth following grazing.

Figure 5.3 Photoassimilate distribution in a rooted cutting of Washington Navel orange (mounted specimen shown on left; matching autoradiograph on right). 14CO2 was supplied to source leaves (boxed area top left) for a day, and movement of 14C-labelled assimilate followed by autoradiography of harvested plant material. 14C photosynthates were distributed widely via vascular conduits to sinks including some roots and a fruit on an adjacent shoot (note stem labelling between sources and sinks). Nearby mature leaves failed to import; they were additional sources of photosynthate. Scale bar = 2 cm (Unpublished material courtesy P.E. Kriedemann)
Photoassimilate transport to harvestable organs plays a central role in crop yield brought about by greater harvest indices. This raises questions about transport and transfer processes that collectively influence photoassimilate partitioning between competing sinks.
Historically, these questions were elucidated by observing partitioning patterns of photoassimilates exported from specified source leaves labelled with 14C supplied as a pulse of 14CO2. Following a chase period, in which 14C photoassimilates are transported to and accumulated by recipient sink organs, the plant is harvested. The pattern of photoassimilate partitioning operating during the pulse is deduced from 14C activity accumulated by sinks (Figure 5.3).
Photoassimilates are partitioned from source leaves to sinks in characteristic and reproducible patterns. For instance, in a vegetative plant, lower leaves are the principal suppliers of photoassimilate to roots, whereas upper leaves are the principal suppliers to the shoot apex. Leaves in an intermediate position export equal quantities of photoassimilates in either direction. However, the pattern of photoassimilate partitioning is not static, it changes with plant development. In vegetative plants, the direction of flow from a leaf changes as more leaves above it become net exporters. Furthermore, at the onset of reproductive development, growing fruits or seeds become dominant shoot sinks for photoassimilates at the expense of vegetative apices.
Photoassimilate partitioning patterns can be altered experimentally by removal of selected sources (e.g. leaves) or sinks (e.g. fruits). These manipulative experiments demonstrate that photoassimilate partitioning reflects the relative strengths of individual sources and sinks. Properties of the phloem pathway connecting sources with sinks are shown in the following Section 5.2.
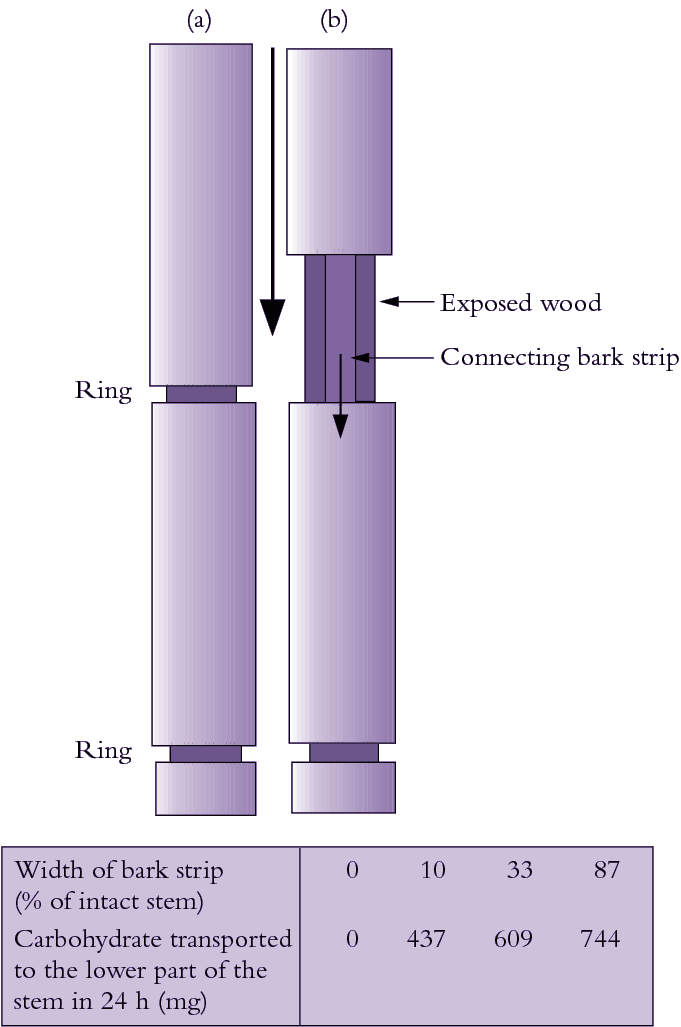
Figure 5.4 The role of bark (phloem) in sugar movement in plants. Mason and Maskell (1928) demonstrated that removing a complete ring of bark (a) while leaving the wood (xylem) intact prevented downward movement of sugars. When a strip of bark was retained between upper and lower stem parts (b), sugars flowed downwards in direct proportion to the width of the remaining bark
Photoassimilate, mainly in the form of sucrose, is loaded into phloem of photosynthetically active leaves for long distance transport to nonphotosynthetic sink tissues. Figure 5.4 shows that assimilate transport occurs in phloem but not xylem. Key characteristics of phloem transport along with its chemical composition and regulation are described below.
In most plant species, phloem is made up of phloem fibres, phloem parenchyma, sieve cells (sieve elements) and their accompanying companion cells (Figure 5.5a). Sieve elements are ideally suited for rapid transport of substances at high rates over long distances. They are elongated and are arranged end to end in files referred to as sieve tubes (Figure 5.16b). Abutting sieve elements are interconnected through membrane-lined pores (sieve pores) with large diameters (1 to 15µm). These pores collectively form sieve plates (Figure 5.16c). The transport capacity of sieve tubes is dependent on a developmentally programmed degeneration of the sieve element protoplasm (cell contents) leaving an open, membrane-bound tube. In mature conducting sieve elements, the protoplast is limited to a functional plasma membrane enclosing a sparse cytoplasm containing low densities of plastids, mitochondria and smooth endo-plasmic reticulum distributed along the lateral walls (Figure 5.16d). These relatively empty sieve tubes provide a longitudinal network which conducts phloem sap (Figure 5.5b).
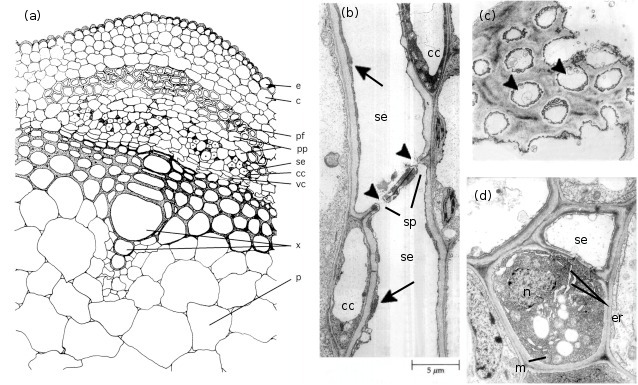
Figure 5.5 (a) spatial arrangement of cell types in a vascular strand from the primary stem of Phaseolus vulgaris (French bean); electron micrographs of stem phloem of Curcurbita maxima (b,c) and P. vulgaris (d) illustrating significant structural characteristics of sieve elements and companion cells. (a) Conducting cells of the phloem (sieve elements) and accompanying companion cells form groups of cells that are separated by phloem parenchyma cells. This mosaic of cells is located between the cortex and xylem and capped by phloem fibres. Scale bar = 7.3 μm. (b) A longitudinal section through two sieve elements arranged end to end to form part of a sieve tube. Companion cells can also be seen. The abutting wall (sieve plates) displays characteristic membrane-lined sieve pores (arrowheads). Cytoplasm of the sieve elements has largely degenerated leaving only endoplasmic reticulum (arrows) and a few plastids around the mature sieve element. Scale bar = 5 μm. (c) A face view of part of a sieve plate showing sieve pores (arrowheads). Scale bar = 0.5 μm. (d) Transverse section through a sieve element and its accompanying companion cell illustrating the sparse cytoplasm and low density of organelles in the sieve element contrasting with the dense ribosome-rich cytoplasm of the nucleated companion cell. Note the mitochondria and rough endoplasmic reticulum. Scale bar = 1.0 μm. c, cortex; cc, companion cell; e, epidermis; er, endoplasmic reticulum; m, mitochondrion; n, nucleus; p, pith; pf, phloem libres; pp, phloem parenchyma; se, sieve element; sp, sieve plates; vc, vascular cambium; x, xylem
Sieve elements are closely associated with one or more companion cells, forming a sieve element–companion cell (se–cc) complex (Figure 5.5d) that plays an important role in transport. These distinct cell types result from division of a common procambial mother cell. In mature se–cc complexes, relatively open sieve elements contrast with adjacent companion cells containing dense, ribosome-rich cytoplasm with a prominent nucleus and abundant mitochondria and rough endoplasmic reticulum (Figure 5.5d). High densities of extensively branched plasmodesmata in contiguous walls of sieve elements and companion cells (Figure 5.6) account for intense intercellular coupling in se–cc complexes (van Bel 1993). Thus, companion cells are considered to perform the metabolic functions surrendered by, but required for, maintenance of viable sieve elements. This functional coupling has led to the concept of se–cc complexes being responsible for phloem transport.
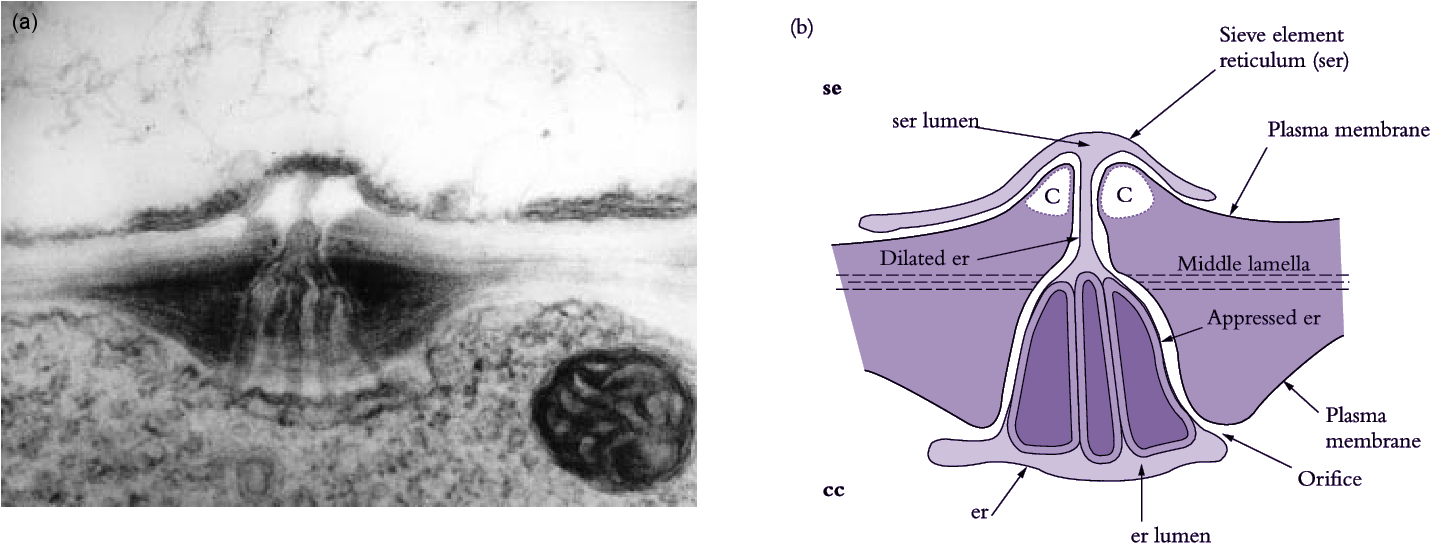
Figure 5.6 (a) Electron micrograph and (b) diagrammatic interpretation of a secondary plasmodesma interconnecting a mature sieve element and its companion cell in a tobacco leaf. Note the characteristic branching of the plasmodesma within the wall of a companion cell. Scale bar = 0.2 μm. c, callose; other symbols as for Figure 5.16 (Based on Ding et al. 1993; reproduced with permission of Blackwell Science)

Figure 5.7 Microautoradiographs of (a) transverse and (b) longitudinal sections of Phaseolus vulgaris stem tissue illustrating localisation of 14C-labelled photosynthate in sieve tubes. These sections are obtained by snap freezing plant tissue and removing frozen water by sublimation (e.g. freeze-drying or freeze substitution). 14C-labelled compounds do not move during preparation. Tissues are embedded in absolute dryness and thin sections are cut, mounted dry on microscope slides and overlain with a thin film of photographic emulsion. Silver grains are visible in the emulsion where 14C, an ideal radioisotope for these experiments, irradiates the film. Abbreviations: se, sieve element; pp, phloem parenchyma; vb, vascular bundle; other symbols as for Figure 5.6. Scale bar in (a) = 20μm; in (b) = 10 μm
Transport of radioactively labelled substances through phloem has been demonstated using microautoradiography (Figure 5.7), providing irrefutable evidence that sieve elements are conduits for transport of phloem sap. Experimentally, a pulse of 14CO2 is fixed photosynthetically and 14C-labelled sugars are given time to reach the stem, which is then excised and processed for microautoradiography. As 14C first moves through the stem, most of the isotope is confined to the transport pathway and very little has had time to move laterally into storage pools. High densities of 14C-labelled sugars are found in sieve elements (Figure 5.7), demonstrating that these cells constitute a transport pathway.
Herbivory or environmental factors causing physical damage could pose a threat to transport through sieve tubes and has undoubtedly imposed strong selection pressure for the evolution of an efficient and rapid sealing mechanism for damaged sieve tubes. Since sieve tube contents are under a high turgor pressure (P), severing would cause phloem contents to surge from the cut site, incurring excessive assimilate loss in the absence of a sealing mechanism. For dicotyledonous species, an abundant phloem-specific protein (P-protein) provides an almost instantaneous seal. P-protein is swept into sieve pores where it becomes entrapped, thus sealing off the damaged sieve tubes. Production of callose (β-1,3 glucan) in response to wounding or high-temperature stress is another strategy to seal off damaged sieve tubes. Callose also seals off sieve pores during overwintering in deciduous plants. Callose is deposited between the plasma membrane and cell wall, eventually blocking sieve pores. Whether deposited in response to damage or overwintering, callose can be degraded by β-1,3 glucanase, allowing sieve tubes to regain transport capacity.
Since phloem translocation is confined to sieve elements embedded within a tissue matrix, it is difficult to obtain uncontaminated samples of translocated sap. The least equivocal approach has been to take advantage of the high P of sieve tube contents. Puncturing or severing sieve tubes should cause exudation of phloem sap provided a sealing mechanism is not activated.

Figure 5.8 Aphids can be used to collect phloem sap. Top photograph: a feeding aphid with its stylet embedded in a sieve tube (see insert); scl, sclerenchyma; st, stylet; x, xylem; p, phloem. Note the drop of ‘honeydew’ being excreted from the aphid’s body. Plates (a) to (e) show a sequence of stylet cutting with an RF microcautery unit at about 3-5 s intervals (a to d) followed by a two-minute interval (d to e) which allowed exudate to accumulate. The stylet has just been cut in (b); droplets of hemolymph (aphid origin) are visible in (b) and (c); once the aphid moves to one side the first exudate appears (d), and within minutes a droplet (e) is available for microanalysis. Scale bars: top = 1 mm, bottom = 1.5 mm (Courtesy D. Fischer)

Figure 5.9. Exudation of phloem contents from lupins. A, following incision of the vasculature at the stylar tip and of the ventral suture of fruits of Lupinus angustifolius. B, following incision of the vasculature of a stem of L. angustifolius. C, following incisions to the vasculature of pre and post anthesis stage flowers on the inflorescence of L. angustifolius. D. exudation at the abscission zone following abscission of two flowers 5 minutes earlier on the raceme of L. mutabilis. Photographs courtesy Craig. Atkins.
For some plant species, sieve-pore sealing develops slowly, or can be experimentally down-regulated by massage or repeated excisions (Milburn and Kallarackal 1989) or slowed by puncturing the vasculature while it is snap frozen in liquid N2 (Pate et al 1984). Carefully placed incisions that do not disturb the underlying xylem, which in any case is more likely to be under tension, permit collection of relatively pure phloem exudate through the severed sieve tubes. Nevertheless, contamination with the contents of cells other than sieve tubes damaged at the site of incision is inevitable. For the major solutes of phloem such as sugars or amino acids that are present in high concentrations this problem is minimal but for less abundant molecules like hormones or other signals, particularly proteins or nucleic acids, conclusions about the origin and functions of these must be made with caution. The ‘natural hemophiliacs’ of the plant world are few and include a number of cucurbits, some brassicas, castor bean, species of the genus Yucca and some species of lupin (Lupinus albus, L. angustifolius, L. mutabilis and L. cosentinii). The excision technique has been expanded to plant species that do not readily exude, by chemically inhibiting the sealing mechanism. Callose production is blocked when wounded surfaces are exposed to the chelating agent ethylenediaminetetraacetic acid (EDTA) by complexing with calcium, a cofactor for callose synthase. Immersing whole, excised organs in EDTA solution, which is essential to inhibit blockage, risks contaminating sap with solutes lost from the apoplast as well as non-conducting cells. This is not an ideal technique.
Enlisting sap-sucking aphids or leaf hoppers to sample sap has been more successful. Aphids can guide a long syringe-like mouthpart (a stylet) into conducting sieve elements (Figure 5.8). Pressure normally forces sieve-tube sap through the stylet into the aphid’s gut where it becomes food or is excreted as ‘honeydew’. By detaching the aphid from its mouthpart pure phloem contents can be collected from the cut end of the implanted stylet. Detaching the aphid body can be achieved by surgery following rapid anesthesia in high CO2 or by severing the stylet using a laser. While stylectomy has been successful with a number of monocotyledons (rice, wheat and barley) the technique has proved more difficult to use with dicotyledons, yielding at best a few microlitres of phloem contents. On the other hand collection of milliliter volumes of exudate from one of the natural hemophiliacs is possible permitting extensive analysis of solutes and macromolecules. In the case of lupins, exudation occurs readily at many sites on the plant so that solutes translocated from source tissues as well as entering sinks can be collected and analysed (Figure 5.9).
Chemical analyses of phloem exudate collected from a wide range of plant species have led to a number of generalisations (e.g. Milburn and Baker 1989) about the contents of sieve tubes. Phloem exudate is a concentrated solution (10–12% dry matter), generating an osmotic pressure (Π) of 1.2 to 1.8MPa. pH is characteristically alkaline (pH 8.0 to 8.5). The principal organic solutes are non-reducing sugars (sucrose), amides (glutamine and asparagine), amino acids (glutamate and aspartate) and organic acids (malate). Of these solutes, non-reducing sugars generally occur in the highest concentrations (300–900 mM). Nitrogen is transported through the phloem as amides and amino acids; nitrate is absent and ammonium only occurs in trace amounts. Calcium, sulphur and iron are scarce in phloem exudate while other inorganic nutrients are present, particularly potassium which is commonly in the range of 60–120 mM. Physiological concentrations of auxins, gibberellins, cytokinins and abscisic acid have been detected in phloem exudate along with nucleotide phosphates. The principal macromolecule group is protein but low levels of peptides and nucleic acids are also present. While in cucurbits the protein in exudate is comprised largely of P-protein, a diverse array of proteins, many of them enzymes, have also been detected.
Phloem sap provides most inorganic and all organic substrates necessary to support plant growth. Non-transpiring tissues are particularly dependent on resources delivered in the phloem (Section 5.1). That translocated sugars represent the major chemical fraction of the phloem sap is consistent with the bulk of plant dry matter (90%) being composed of carbon, hydrogen and oxygen. Carbon transport is further augmented by transport of nitrogen in organic forms.
Carbohydrate is translocated as non-reducing sugars in which the metabolically reactive aldehyde or ketone group is reduced to an alcohol (mannitol, sorbitol) or combined with a similar group from another sugar to form an oligosaccharide. Apart from sucrose, transported oligosaccharides belong to the raffinose series. In this series, sucrose is bound with increasing numbers of galactose residues to form raffinose, stachyose and verbascose respectively. However, sucrose is the most common sugar species transported. In a small number of plant families, other sugar species predominate. For example, the sugar alcohol sorbitol is the principal transport sugar in the Rosaceae (e.g. apple) and stachyose predominates in the Cucurbitaceae (e.g. pumpkin and squash). Exclusive transport of non-reducing sugars probably reflects packaging of carbohydrate in a chemical form which protects it from being metabolised. Metabolism of these transported sugars requires their conversion to an aldehyde or ketone by enzymes which are thought to be absent from sieve-tube contents.
Plant physiologists have long regarded the two long distance translocation streams of xylem and phloem as having functions additional to the distribution of nutrients and assimilates. Specifically, each serves as a means of communication between the source and sink organs such that systemic signals are thought to transmit molecular responses to endogenous and environmental cues. Furthermore, evidence is accumulating that some of these signals regulate gene expression as a consequence of their translocation (see below).
(c) Macromolecule composition of phloem
Proteomic and transcriptomic analyses have demonstrated a widely diverse composition of proteins, peptides and nucleic acids, including mRNA and small RNAs, in phloem exudates. While the origin of each individual protein or nucleic acid remains to be verified the limited compositional data available from stylectomy confirms that indeed each group of macromolecules is present in phloem. In cucurbit phloem exudate some 1110 different proteins have been detected along with a large number of mRNAs and similar data have been obtained for exudates from other species (Brassica napus, Ricinus communis and Lupinus albus). Compositional data for phloem proteins of these species show a common complement that includes phloem-specific P proteins together with proteins involved in sugar metabolism and transport, protein turnover and transport, detoxification of reactive oxygen species, as well as proteins that provide defence against insect herbivores and pathogens (Figure 10). Some undoubtedly play a role in maintenance of the SE system while others, such as the Flowering Locus T (FT) protein associated with the flowering response (‘florigen’), appear to be systemic ‘signals’ (Rodriguez-Medina et al 2011) and there may be many more. Because sieve tubes are enucleate and lack ribosomes (5.2.2 a), proteins in the translocation stream are not formed in situ but are transported from sites of synthesis in the companion cells.

Figure 5.10. Two dimensional polyacrylamide gel electrophoresis separation of proteins in phloem exudate from Lupinus albus. The gel was developed in the first dimension by isoelectric focusing with a linear pH gradient of 3-10 followed by separation due to differences in molecular mass. The positions of mass standards are shown on the right hand side of the gel. After staining with Coomassie Blue to locate the spots they were excised for digestion with trypsin. The peptides were then analysed by partial sequence determination using MS/MS and identified using database searches. (Courtesy of Craig Atkins)
Functional analysis of the cDNA identified in transcriptome studies of phloem exudates revealed transcripts involved in a wide range of processes that include metabolism, plant responses to stresses, transport, DNA/RNA binding and protein turnover. The presence of transcripts in phloem exudate supports the idea of an RNA-based signalling network that is thought to function in control of processes associated with plant growth and development (Lough and Lucas, 2006). However, the functional role of transcripts in the contents of sieve tubes as well as their actual translocation is yet to be determined.
Small RNA molecules (18-25 nt) have been identified in phloem exudate collected from rape, white lupin, pumpkin, castor bean and Yucca filamentosa as well as in aphid stylet exudate collected from apple stems. The population includes both microRNAs (miRNA) and small interfering RNAs (siRNA) a large number of which target mRNA of transcription factors that themselves regulate genes expressions. miRNAs are also involved in mediating environmental responses, including responses to salinity, drought, nutrient limitations, as well as hormone interactions. Their small size and powerful functions in targeting mRNAs to regulate expression suggest that those in phloem exudate are likely to be systemic signals.
An important question that relates to the significance of macromolecules in the contents of sieve tubes is proof that they are translocated and that translocation is essential for their function at a sink. A diversity of studies that have exploited cucurbit root stocks and grafted scions has provided clear evidence that P proteins among others are graft transmissible. In a series of elegant experiments Aoki et al. (2005) labelled and injected two isolated pumpkin phloem proteins (CmPP16‑1 and CmPP16-2) into the vasculature of intact rice plants through severed leaf hopper stylets and showed their translocation as well as some evidence for specificity in protein translocation. The Flowering Locus T (FT) protein formed in leaves mediates the flowering transition of shoot apical meristems and the evidence that it is translocated is compelling. The long distance movement of RNA molecules was first demonstrated for plant viruses and there is now good evidence for phloem translocation of a number of transcripts (Lough and Lucas 2006). A recent compilation identified 13 miRNAs involved in plant responses to drought/salt stress (Covarrubias and Reyes 2010). Eight of these were identified in lupin phloem exudate (Rodriguez-Medina et al. 2011) and, importantly, six were also recovered from PCR amplification of apple stylet exudate (Varkonyi-Gasic et al 2010). There is thus a possibility that the responses to drought and salinity are mediated through miRNAs translocated from sites where the stress is sensed to sites where a response is initiated.
The most convincing case for a translocated miRNA in phloem regulating gene expression relates to Pi homeostasis. While both local and systemic signals are involved, miR399 is phloem mobile and acts directly in roots to down regulate the expression of PHO2 (a ubiquitin conjugating enzyme) that results in greater expression of Pi transporters to increase Pi uptake under conditions of deficiency. Systemic signaling has also been implicated in homeostasis of other nutrients, including N, S and Cu with, in each case, miRNAs involved.
Phloem flux can be estimated in a number of ways. The simplest is to determine dry weight gain of a discrete organ connected to the remainder of a plant by a clearly definable axis of known phloem cross-sectional area. Developing fruits or tubers meet these criteria. Sequential harvests from a population of growing fruit or tubers provide measures of the organ’s net gain of dry matter imported through the phloem. Net gains or losses of dry matter resulting from respiration or photosynthesis are incorporated into calculations to give gross gain in dry matter by the organ. Flux of dry matter through the phloem (specific mass transfer — SMT; Canny 1973) can then be computed on a phloem or preferably on a sieve-tube lumen cross-sectional area basis. Area estimates can be obtained from histological sections of the pedicel or stolon that connects a test organ to its parent plant. Expressed on a phloem cross-sectional area basis, SMT estimates are normally in the range of 2.8–11.1 g m–2 phloem s–1 (Canny 1973). Flux on the basis of sieve-tube lumen cross-sectional area is preferable but relies on identification of sieve tubes and the assumption that they are equally functional as transport conduits. Sieve tubes account for some 20% of phloem cross-sectional area, suggesting fluxes are about five-fold higher through a sieve-tube lumen.
Speed of phloem translocation can be determined from simultaneous measurements of SMT and phloem sap concentrations as shown in Equation 5.1 below:
\[\mathrm{Speed} (m \cdot s^{-1}) = \mathrm{SMT}(g \cdot m^{-2} \cdot s^{-1})/ \mathrm{concentration} (g \cdot m^{-3}) \tag{5.1} \]
For a sucrose concentration of 600 mM (or 2.16 x 105 g m-3) and the highest SMT values shown above, Equation 5.1 estimates that phloem sap can move at speeds of up to 56 × 10–5 m s–1 or 200 cm h–1. These estimates have been verified by following the movement of radioisotopes introduced into the phloem translocation stream.
These estimates of transport rates and speeds tacitly assume that phloem sap moves through sieve tubes by mass flow (water and dissolved substances travel at the same speed). Independent estimates of transport rate, concentration of phloem sap and translocation speed lend support to, but do not verify, the assumption that movement occurs as a mass flow.
A simple and direct test for mass flow is to determine experimentally whether water and dissolved substances move at the same speed. This test should be relatively easy to apply using radioactively labelled molecules. Unfortunately, in practice it turns out that different molecular species are not loaded into the sieve tubes at the same rates and the plasma membranes lining the sieve tubes are not equally permeable to each substance. Thus, the analysis is complicated by the necessity to use model-based corrections for rates of loading into and losses from the sieve tubes. Nevertheless, the speed estimates obtained from such experiments are found to be similar for dissimilar molecules, supporting the proposition that mass flow accounts for most transport through sieve tubes.
Phloem translocation is generally believed to be driven by pressure. Münch (1930) proposed that a passive mass flow of phloem sap through sieve tubes was driven by the osmotically generated pressure gradient between source and sink regions (Figure 5.11). At source regions, the principal osmotica of phloem sap are actively or passively loaded into sieve tubes from companion cells or mesophyll cells (see 5.3.2), thereby driving water towards the lower water potentials within sieve tubes. As water enters, P rises. Unloading of solutes from sieve tubes at sink regions reverses water potentials; water flows out of sieve tubes and P falls relative to that of sieve tubes in source regions.
The pressure-flow hypothesis can be modelled using the relationship that rate of mass flow (Ff) of a substance is given by the product of speed (S) of solution flow, path cross-sectional area (A) and its concentration (C). That is:
\[F_f = S \cdot A \cdot C \tag{5.2}\]
Speed (m s–1) has the same units as volume flux (Jv — m3 m–2 s–1) of solution passing through a transport conduit. Poisseuille’s Law describes the volume flux (Jv) of a solution of a known viscosity (h) driven by a pressure difference (DP) applied over the length (l) of pathway of radius (r) as:
\[J_v = \pi r^4 \Delta P/8 \eta l \tag{5.3} \]

Figure 5.11 Scheme describing the pressure flow hypothesis of phloem transport (Based on Münch 1930)
The term πr4/8ηl in Equation 5.3 provides an estimate of hydraulic conductivity (Lp) of the sieve-tube conduit which is set by the radius of the sieve pores. Raised to the fourth power, small changes in the sieve-pore radius will exert profound effects on the hydraulic conductivity of the sieve tubes (Section 5.2). The viscosity of sieve-tube sap is determined by the chemical species (particularly sugars) and their concentrations in the phloem sap.
Key features of the pressure-flow hypothesis are encapsulated in Equation 5.3. The central question is whether a pressure gradient exists in sieve tubes with the expected direction and of sufficient magnitude to support observed rates of sap flow. Indirect estimates of P in sieve tubes made through determination of intra- and extracellular P support the pressure-flow hypothesis. Direct measurements of sieve-tube P are technically challenging because of the inaccessibility of these small, highly turgid cells. They are, for instance, too small for pressure-probe measurements. However, manometric pressure measurements obtained using severed aphid stylets agree with indirect estimates (Wright and Fisher 1980). Experimental manipulation of the pressure gradient between the source and sink also results in alterations in phloem translocation rates consistent with the pressure-flow model.
Whether the pressure gradient is sufficiently steep is a more vexing question. The pressure gradient required to drive phloem translocation at observed rates is determined by the transport resistance of the phloem path, according to Ohm’s Law. Dimensions of the sieve pores set a limiting radius for volume flux of transported sap (Equation 5.3) and hence transport resistance. If the sieve pores were open and unoccluded by P-protein, a number of studies have demon-strated that the measured pressure gradients are sufficient to support the observed rates of flow. However, the in situ radii of sieve pores remain unknown.
Overall, the pressure-flow hypothesis accounts for many observed features of phloem translocation, including distribution of resources. While conclusive evidence supporting this hypothesis is still sought, less attention is now focused on this issue with a growing appreciation that the phloem pathway has spare transport capacity. Evidence from Kallarackal and Milburn (1984), for example, showed that the specific mass transfer (SMT – see preceding section) to an intact fruit of castor bean could be doubled on removal of competing fruits. Moreover, if P of sieve elements at the sink end of the phloem path was reduced to zero, by severing the pedicel and allowing exudation, SMT rose to an incredible 305 g m–2 sieve-tube area s–1! In another experiment, when half the conducting tissue was removed from the peduncle of sorghum or wheat plants, grain growth rate was not impaired (Wardlaw 1990). Together, these observations imply that phloem has excess carrying capacity in both dicotyledons and monocotyledons. Particularly in monocotyledonous plants, a strong selection pressure for spare transport capacity must exist because there is no vascular cambial activity to replace damaged sieve elements.
Loading of sugars, potassium and accompanying anions into sieve tubes at sources determines solute concentrations in phloem sap (Table 5.1). The osmotic pressure (Π) of these solutes influences P generated in sieve tubes. Thus, source output determines the total amount of assimilate available for phloem transport as well as the pressure head driving transport along the phloem path to recipient sinks. Withdrawal of assimilates from sieve tubes at the sink end of the phloem path, by the combined activities of phloem unloading and metabolism/compartmentation (Table 5.1), determines Π of phloem sap. Other sink-located membrane transport processes influence Π around sieve tubes. The difference between intra- and extracellular Π of sieve tubes is a characteristic property of each sink and determines P in sink sieve tubes.
The pressure difference between source and sink ends of the phloem pathway drives sap flow (Equation 5.3) and hence phloem translocation rate (Equation 5.2) from source to sink. The source and sink processes governing the pressure dif-ference (Table 5.1) are metabolically dependent, thus rendering phloem translocation rates susceptible to cellular and environmental influences. The pressure-flow hypothesis predicts that the phloem path contribution to longitudinal transport is determined by the structural properties of sieve tubes (Table 5.1). Variables of particular importance are cross-sectional area (A) of the path (determined by numbers of sieve pores in a sieve plate and sieve-tube numbers) and radius of these pores (sets r in Equation 5.3). These quantities appear in Equations 5.2 and 5.3. Thus, the individual properties of each sink and those of the phloem path connecting that sink to its source will determine the potential rate of assimilate import to the sink (Figure 5.12).
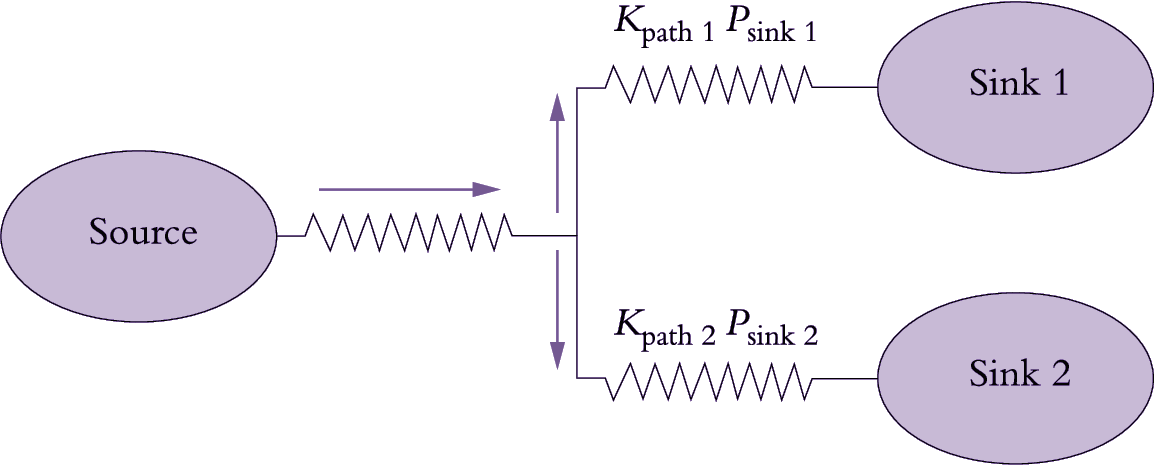
Figure 5.12. Scheme describing photoassimilate flow from a source leaf linked to two competing sinks, Sink 1 and Sink 2. Assimilate flows through alternative phloem paths (Path 1 and Path 2) each with its own conductance (Kpath) and pressure difference (P) between source and sink. Hence Path 1 is distinguished by Kpath1 and Psink1 and Path 2 by Kpath2and Psink2
The transport rate (R) of assimilate along each phloem path, linking a source with each respective sink, can be predicted from the pressure-flow hypothesis (see Equations 5.2 and 5.3) as:
\[ R = K_{path} (P_{source} - P_{sink}) C \tag{5.4}\]
where path conductance (Kpath) is the product of path hydraulic conductivity (Lp) and cross-sectional area (A). Hence, the relative flows of assimilates between hypothetical sinks (sink 1 and sink 2) shown in Figure 5.12 may be expressed by the following ratio:
\[ \frac{K_{path1} (P_{source} - P_{sink1}) C} {K_{path2} (P_{source} - P_{sink2}) C} \tag {5.5} \]
Partitioning of assimilates between two competing sinks is thus a function of path conductance and P at the sink end of the phloem path (Equation 5.5). Since phloem has spare capacity, any differences in the conductance of the inter-connecting paths (Figure 5.12) would exert little influence on the rate of phloem transport to the competing sinks. Assimilate partitioning between competing sinks would then be determined by the relative capacity of each sink to depress sieve-tube P at the sink end of the respective phloem path. Even when differences in path conductance are experi-mentally imposed, phloem transport rates are sustained by adjustments to the pressure differences between the source and sink ends of the phloem path (Wardlaw 1990).
These conclusions have led to a shift in focus from phloem transport to phloem loading and unloading, which are instrumental in determining the amount of assimilate translocated and its partitioning between competing sinks, respectively.
Photoassimilates are loaded along the entire phloem transport pathway, from photosynthetic leaves to importing sinks. While most photoassimilate loading occurs in photosynthetically active leaves, root-produced metabolites, such as amino acids, move readily from xylem to phloem particularly at the stem nodes. Phloem loading also occurs in storage organs during periods when reserves are remobilised and exported. Indeed, the membrane transport events contributing to phloem loading were first examined using export of sucrose remobilised from the endosperm of germinating castor bean seed as an experimental model (Kriedemann and Beevers 1967).
This section focuses on phloem loading in the leaves. It analyses the cellular pathways for assimilate loading, and the regulatory controls.
Phloem loading is used variously to describe transport events outside, and inside, phloem tissues of leaves. The broader general application is adopted here — that is, phloem loading describes photoassimilate transport from the cytoplasm of photosynthetic mesophyll cells to se–cc complexes of leaf phloem.
Phloem loading commences in mesophyll cells and ends in the leaf vascular system. The se–cc complexes occur in a wide array of vascular bundle sizes. In dicotyledonous leaves, veins undergo repeated branching, forming the extensive minor vein network described in Section 5.2. For example, sugar beet leaves contain 70cm of minor veins cm–2 of leaf blade, while the major veins contribute only 5.5cm cm–2 of leaf blade (Geiger 1975). These observations and physiological studies (van Bel 1993) show that the principal site of phloem loading is in the minor vein network of dicotyledonous leaves. In contrast, the major veins transport loaded photoassimilates out of leaves.
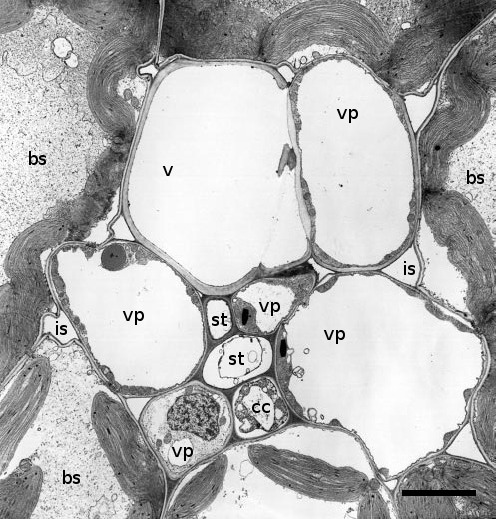
Figure 5.13. Transmission electron micrograph through a minor vein of a source leaf of maize (Zea mays L.) This vascular bundle consists of two sieve elements (st), one xylem vessel (v) and five vascular parenchuma cells (vp). These sieve elements are of two types, one thin walled and accompanied by a companion cell, the other thick walled and adjacent to the xylem vessel. Other symbols are: bs, bundle sheath; cc, companion cell; is, intracellular space; st, sieve tube. Scale bar - 4.2 µm (Based on Evert et al. 1978; reproduced with permission of Planta)
Minor veins usually comprise a single xylem element, vascular parenchyma cells and one to two sieve elements surrounded by one to four companion cells (Figure 5.13). The se–cc complex in minor veins bears similarities to that of stems (Figure 5.6). Companion cells have dense cytoplasm containing many mitochondria and are often considerably larger than the sieve elements they accompany. Companion cells are symplasmically connected to the sieve elements by branched plasmodesmata.
Cross-sectional areas of veins in monocotyledonous leaves reveal large and small parallel veins. Photoassimilates are loaded into the small veins and conducted through large veins. Fine transverse veins carry photoassimilates loaded into small veins across to large veins for export.
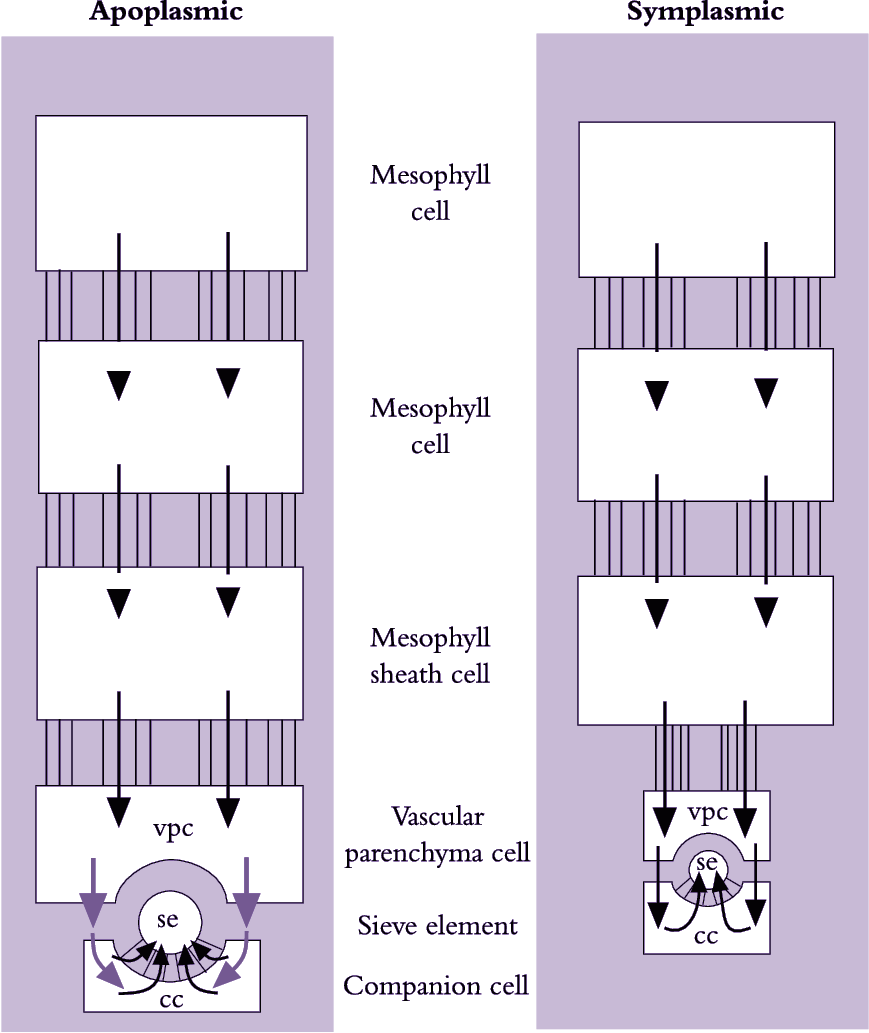
Figure 5.14. Scheme describing symplasmic and apoplasmic pathways of phloem loading. Lines without arrows joining boxes represent symplasmic continuity (i.e. plasmodesmata). Black arrows indicate symplasmic transport (i.e. through plasmdesmata); green arrows indicate apoplasmic transport requiring solutes to cross membranes. vpc, vascular parenchyma cell; se, sieve element; cc, companion cell (Based on van Bel 1993)
Photoassimilates could move intercellularly through interconnecting plasmodesmata from chloroplasts in mesophyll cells to the lumena of sieve elements (symplasmic phloem loading) or across plasma membranes, travelling part of the route through the cell wall continuum (apoplasmic phloem loading). These fundamentally different pathways are shown schematically in Figure 5.14. Debate persists over which cellular pathway of phloem loading prevails because experiments on transport from mesophyll cells to sieve elements are difficult.
Extraordinarily, the cellular pathway of phloem loading reflects evolutionary relationships. Species from ancient plant groups display symplasmic loading, while species of more modern plant groups appears to exhibit apoplasmic phloem loading (van Bel 1993). Evidence for respective routes of loading follows.
A symplasmic pathway depends upon development of extensive plasmodesmal interconnections between adjoining cells, forming a cytoplasmic continuum from mesophyll to se–cc complexes (Figure 5.14). Such symplastic continuity is found in leaves of plant families containing trees and shrubs as well as cucurbits such as squash (van Bel 1993). An abundance of plasmodesmal interconnections demonstrates potential for symplasmic transport but does not establish whether such transport actually occurs. Membrane-impermeant fluorescent dyes microinjected into mesophyll cells are transported to se–cc complexes, demonstrating that plasmodesmata can provide a route for photoassimilate transport. Furthermore, when leaves were fed14CO2 and treated with inhibitors that block sugar transport across plasma membranes, transport of 14C-labelled photo-assimilates continued unaffected along the enforced symplasmic unloading route (Figure 5.15; van Bel 1993). In this case, sugar levels are higher in the mesophyll than in the phloem and ions and molecules diffuse through plasmodesmata at each interface, without a concentrating step (Turgeon 2010). Therefore, this is a passive symplasmic phloem loading.
Symplasmic phloem loading may also be an active process occurring in some herbaceous eudicots. This model of phloem loading, called polymer trap mechanism, depends on sucrose being biochemically converted to raffinose oligosaccharides (RFOs) in specialized CCs (intermediary cells - ICs) (Turgeon 2010). The biochemical synthesis of RFOs from sucrose requires metabolic energy. The synthesized RFOs exceed size exclusion limits of plasmodesmata linking mesophyll cells with ICs and therefore are trapped and accumulate to high concentrations in SE/IC complexes of minor veins for long distance transport (Turgeon 2010).
Plant species that load phloem from the leaf apoplasm are characterised by a low abundance of plasmodesmata between se–cc complexes and abutting vascular cells. However, as for symplasmic loaders, mesophyll cells of these species are interconnected by abundant plasmodesmata (Figure 5.23). Herbaceous and many crop species belong to this group of phloem loaders, including grasses (van Bel 1993). Conventional physiological observations are consistent with phloem loading in leaves of these species including a membrane transport event located somewhere between mesophyll cells and the se–cc complexes of minor veins (Figure 5.15).
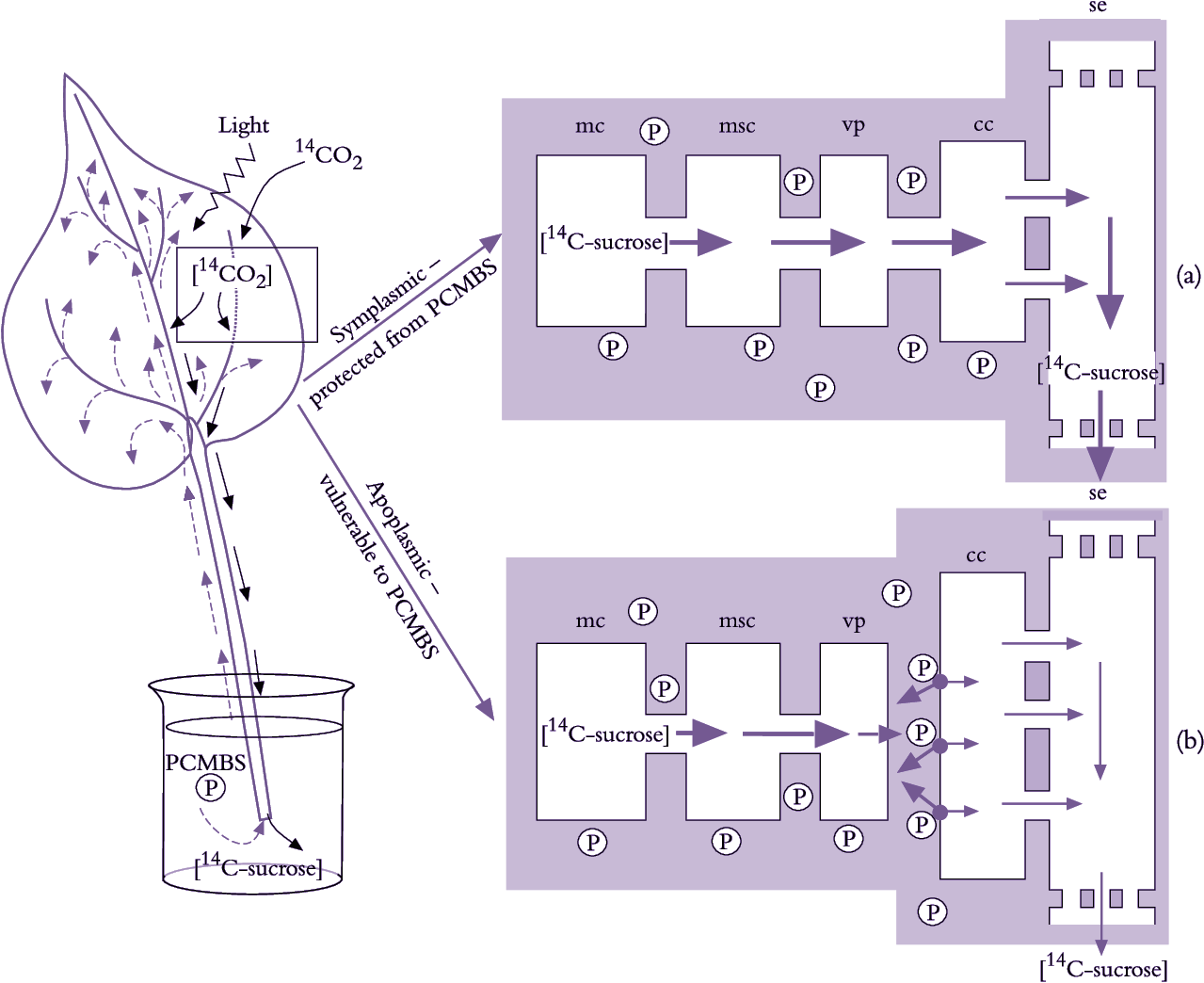
Figure 5.15. Testing whether photoassimilates move from mesophyll cells to se-cc complexes through (a) an entirely symplasmic route or (b) a route with an apoplasmic step. The approach is to use PCMBS as an inhibitor of membrane transport. PCMBS does not cross membranes but binds to the apoplasmic face of plasma membranes. Therefore, it blocks apoplasmic transport while symplasmic phloem loading is unaffected. PCMBS was introduced into the leaf apoplasm through the transpiration stream of excised leaves. Leaves were then exposed in a closed illuminated chamber to 14CO2. The 14C photoassimilate exported from labelled leaf blades was used to monitor phloem loading. PCMBS only reduced photoassimilate export (i.e. phloem loading) from those leaves with few plasmodesmata interconnecting se-cc complexes with surrounding cells. Thus, photoassimilate flow included a membrane transport step from the leaf apoplasm in certain plant species while others loaded via a symplasmic route. cc, companion cell; mc, mesophyll cell; msc, mesophyll sheath cell; PCMBS (para-chloromercuriben-zenesulphonic acid, also abbreviated to P); se, sieve element; vp, vascular parenchyma (Based on van Bel 1993)
Molecular biology has brought new insights to phloem loading. For instance, existence of an apoplasmic step demonstrated with PCMBS (Figure 5.15) has been elegantly confirmed using molecular biology to control activity of the sucrose/proton symporter responsible for sucrose uptake from phloem apoplasm into se–cc complexes. Specifically, potato plants were transformed with an antisense copy of the gene encoding the sucrose/proton symporter, producing a phenotype with low levels of the symporter in plasma membranes of se–cc complexes (Frommer et al. 1996). Excised leaves of transformed plants exported significantly less photoassimilates than wild-type plants, corroborating the inhibitory effect of PCMBS on apoplasmic phloem loading (Figure 5.15). This provides compelling evidence that passage of photoassimilates from mesophyll cells to se–cc complexes in potato leaves includes an apoplasmic step.
Vascular parenchyma cells are the most probable site for photoassimilate exchange to phloem apoplasm (van Bel 1993), ensuring direct delivery for loading into se–cc complexes. Furthermore, plasma membranes of se–cc complexes in minor veins have increased surface areas to support photoassimilate transfer from phloem apoplasm. Notably, the surface area of se–cc complexes in sugar beet leaves is surprisingly large—0.88 cm–2of leaf blade surface. By implication, these large membrane surfaces are involved in phloem loading. Further support comes from cytochemical studies, demonstrating a great abundance of proteins associated with energy-coupled sucrose transport (Section 5.3.3(b)).
Leaf anatomies in some plant species suggest a potential for simultaneous phloem loading through apoplasmic and symplasmic pathways (van Bel 1993). Whether these pathways connect the same sieve element, different sieve elements in the same minor vein order or sieve elements in different vein orders is still unknown.
Any hypothesis of phloem loading must account for the following characteristics:
The above-described characteristics have been used to argue against loading of se–cc complexes through a symplasmic route on the grounds that plasmodesmata lack mechanisms for concentrating and selecting solutes. However, a contribution of plasmodesmata to concentrating and selecting solutes cannot be precluded from our current knowledge of plasmodesmal structure and function.
Plants that load se–cc complexes through a symplasmic route translocate 20–80% of sugars in the form of raffinose-related compounds such as raffinose, stachyose and verbascose (Section 5.2.3(c)). Grusak et al. (1996) proposed a model for symplasmic phloem loading that accounts for the general characteristics stated above. According to this model (Figure 5.16), sucrose diffuses from mesophyll and bundle sheath cells into intermediary (companion) cells through plasmodesmata. Within companion cells, sucrose is thought to be enzymatically converted to oligosaccharides (raffinose or stachyose) maintaining a diffusion gradient for sucrose from mesophyll cells into se–cc complexes. The molecular-size-exclusion limit of plasmodesmata interconnecting mesophyll and companion cells is such that it prevents back diffusion of stachyose and raffinose molecules, which are larger than sucrose. These oligosaccharides are able to diffuse through plasmodesmata with larger diameters linking companion cells with sieve elements (van Bel 1993). This model accounts for selective loading of sugars to achieve high photoassimilate concentrations in phloem elements.
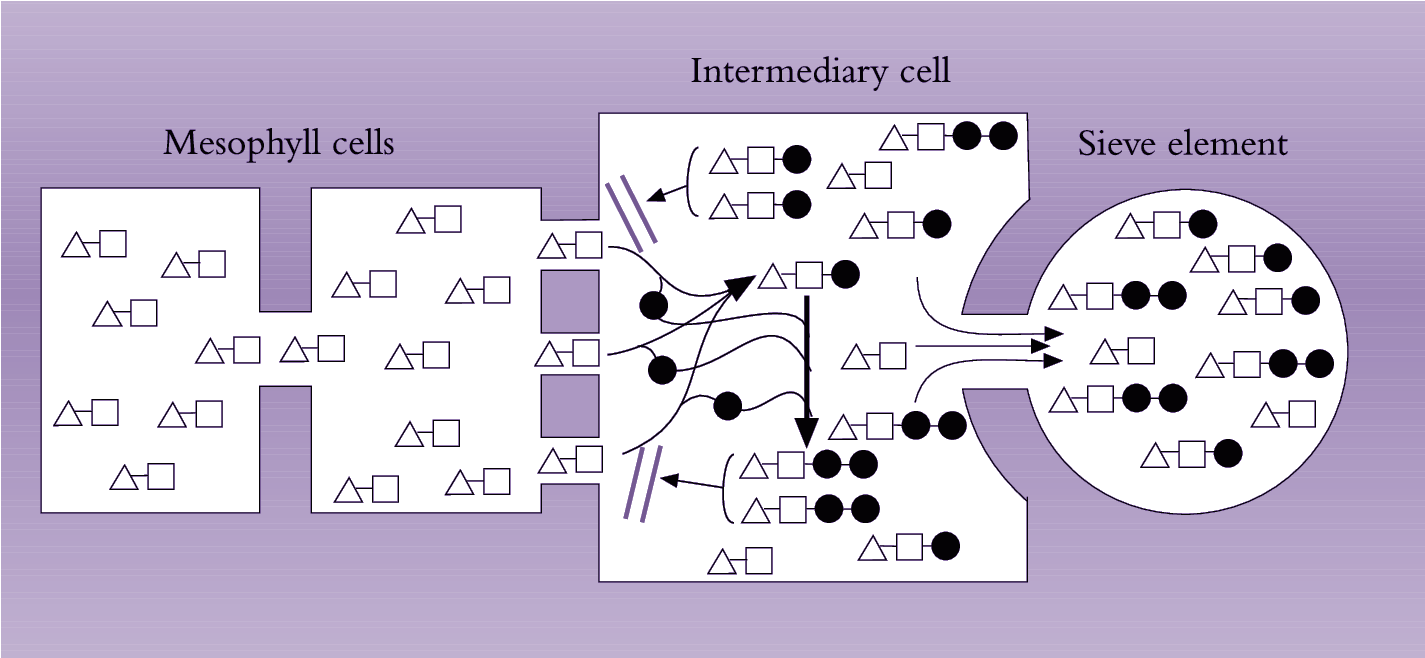
Figure 5.16. Model of the ‘polymerisation trap mechanism’ to explain symplasinic phloem loading against a solute concentration gradient. Sucrose moves through a symplasmic path from photosynthetic cells into intermediary (companion) cells of the minor veins. Sucrose movement is by diffusion down a concentration gradient maintained by the polymerisation of sucrose into oligosaccharides (raffinose and stachyose) in intermediary cells. Diffusion of these oligosaccharides into mesophyll cells is prevented, as their size exceeds the molecular exclusion limit of plasmodesmata joining mesophyll and intermediary cells. However, the larger-diametered plasmodesmata linking intermediary cells with sieve elements permit oligosaccharides to be loaded into sieve elements for export from the leaf. [], glucose; Δ, fructose; • galactinol (Based on Grusak et al. 1996)
Phloem loading with an apoplasmic step is an attractive model, explaining both how solutes become concentrated in se–cc complexes (energy-coupled membrane transport) and how they could be selected by specific membrane transporters (see van Bel 1993). Identifying transport mechanisms responsible for photoassimilate transport to and from the leaf apoplasm has proved challenging.
Based on estimates of sucrose fluxes and high sucrose concentrations in phloem sap, there is little doubt that sucrose loading into phloem is energy dependent. The demonstration that PCMBS blocks loading of photoassimilates in whole leaves of certain species (Section 5.3.2(b)) points to carrier-mediated transport across plasma membranes. Genes encoding sucrose porters have been cloned from leaf tissue (Frommer et al. 1996) and shown to be specifically expressed in leaf phloem. Complementation studies in yeast defective in sucrose transport suggest that the phloem-located sucrose porter catalyses sucrose/proton symport in a similar way to that illustrated in Figure 5.32. Antisense transformants of potato with low abundance of this symporter have impaired sucrose transport (Section 5.32(b)).
In contrast to photoassimilate uptake from phloem apoplasm, very little is known about the mechanism of sugar efflux into the apoplasm until very recently. Estimates of photoassimilate flux to phloem apoplasm, based on rates of sucrose export from leaves, suggest that this transport event must be facilitated by other transport processes (van Bel 1993). This is now confirmed by the recent cloning of sucrose efflux protein that sheds a light on the molecular mechanisms of phloem loading (Chen et al. 2012).

Figure 5.17. Time-course of photoassimilate export from source leaves of tomato plants. Control plants, in which fruits were a major sink for photoassimilates, were maintained at 20°C. Treatments involved (1) removing fruit or (2) exposing plants with fruits to 30°C. The proportion of 14C label remaining in source leaves after a radioactive pulse was monitored through time to show that (1) presence of major sinks or (2) more rapid metabolism accelerated 14C export from source leaves (Based on Moorby and Jarman 1975).
The response of photoassimilate export to changes in sink demand depends upon whether photoassimilate flow is source or sink limited (Wardlaw 1990). A source-limited system does not respond rapidly to an increase in sink demand, depending more on the capacity of leaves to increase the size of the transport pool. In contrast, alterations in sink demand in a sink-limited system elicit immediate effects on photoassimilate export. Figure 5.17 shows how the presence of fruits accelerates 14C export, especially at high temperatures. For leaves that load the se–cc complexes from apoplasmic pools, changes in sink demand probably influence photoassimilate export by altering membrane transport properties. These changes in membrane transport entrain a flow of adjustments in biochemical partitioning within the leaf through substrate feedback (see below).
Changes in the turgor pressure of phloem sap or altered phytohormone levels could serve as signals for sink demand.
Changes in the pressure of sink phloem sap are rapidly transmitted through sieve tubes to sources. Phloem loading in source tissues responds to this pressure signal by changes in solute transport rates mediated by membrane-associated porters (van Bel 1993). This is a proposed mechanism for phloem loading which would respond rapidly (within minutes) to changes in sink demand.
Phytohormone levels in leaves respond to changes in the source/sink ratio. For instance, gibberellin levels in leaves proximal to developing inflorescences increase at fruit set. In contrast, abscisic acid levels in soybean and grape leaves are inversely related to alterations in sink demand (Brenner 1987). Therefore, changes in leaf phytohormone levels could serve to signal shifts in sink demand for photoassimilates. In this context, direct application of auxin and gibberellic acid to source leaves results in a rapid enhancement of photoassimilate export (Table 5.2). Gibberellic acid did not stimulate leaf photosynthesis or alter photoassimilate partitioning, appearing instead to upregulate phloem loading. This was confirmed by faster 14C loading into isolated phloem strands (Table 5.2).
A substrate feedback response is elicited if the rate of photo-assimilate export from chloroplasts is limited by sink demand. If sucrose export from source pools is accelerated by phloem loading, substrate feedback inhibition of photoassimilate delivery is alleviated. A cascade of adjustments in the activity of key regulatory enzymes follows (see Section 2.3) with the final outcome of an increased flow of sucrose into transport pools. Conversely, if photoassimilate flow is limited by photosynthetic rate, the activity of enzymes responsible for sucrose biosynthesis is not subject to feedback inhibition by substrates. As a consequence, responses to increased sink demand can only be mediated by increases in photosynthetic enzyme activity.
Photoassimilate removal from phloem and delivery to recipient sink cells (phloem unloading) is the final step in photoassimilate transport from source to sink. Within sink cells, cellular metabolism and compartmentation are the end-users of phloem-imported photoassimilates. Combined activities of these sink-located transport and transfer events determine the pattern of photoassimilate partitioning between competing sinks and hence contribute to crop yield.
Phloem unloading describes transport events responsible for assimilate movement from se–cc complexes to recipient sink cells. A distinction must be made between transport across the se–cc complex boundary and subsequent movement to recipient sink cells. The former transport event is termed sieve element unloadingand the latter post-sieve element transport. On reaching the cytoplasm of recipient sink cells, imported photoassimilates can enter metabolic pathways or be compartmented into organelles (e.g. amyloplasts, protein bodies and vacuoles). Metabolic fates for photoassimilates include catabolism in respiratory pathways, biosynthesis (maintenance and growth) and storage as macromolecules (starch and fructans).
Compared with phloem loading, phloem unloading and subsequent sink utilisation of imported photoassimilates operate within a much broader range of configurations:
A correspondingly large range of strategies for phloem unloading and sink utilisation must be anticipated.
Most photoassimilates travel along one of three cellular pathways: apoplasmic, symplasmic or a combination of both with symplasmic transport interrupted by an apoplasmic step (Figure 5.19).
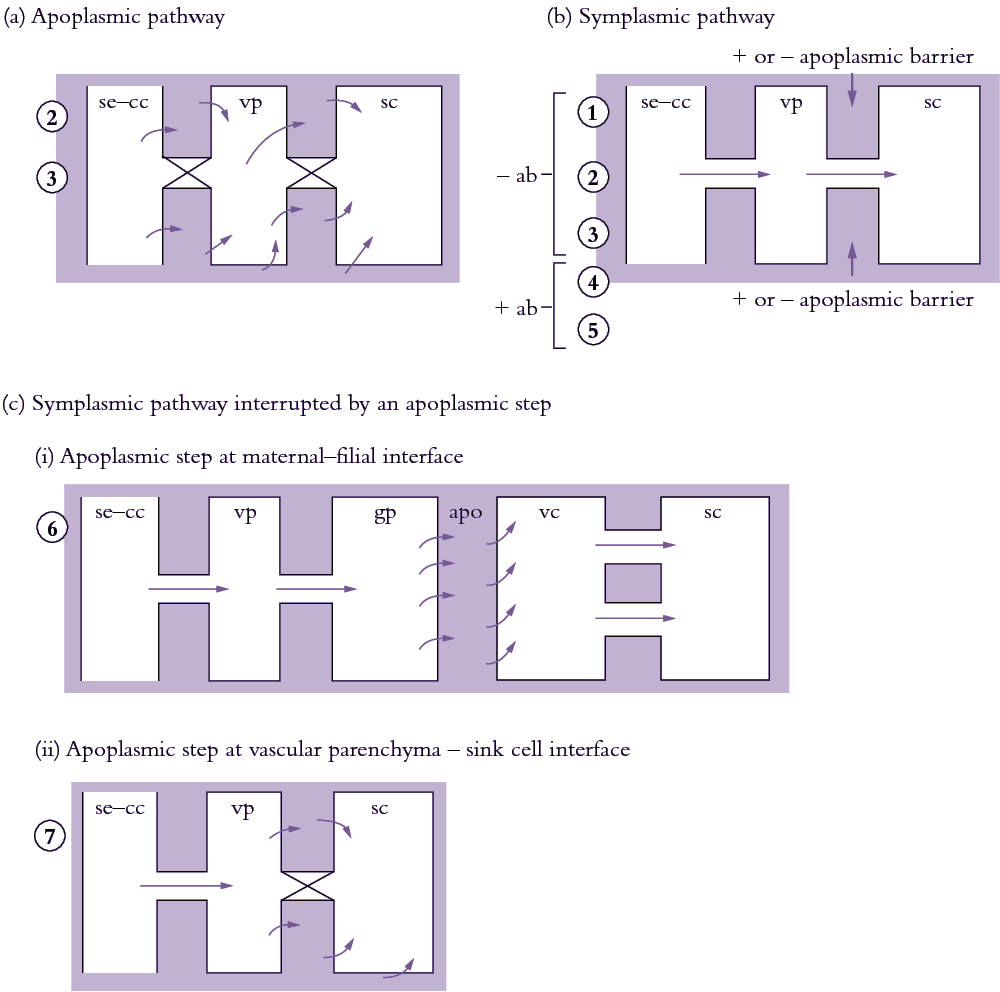
Figure 5.18. Scheme describing the cellular pathways of phloem unloading and their relationship with sink types. (a) Apoplasmic unloading showing direct transport of photoassimilates from se—cc complexes to the phloem apoplasm. (b) Symplasmic unloading pathway which may or may not have an apoplasmic barrier between sieve elements and recipient sink cells. (c) Symplasmic unloading with the intervention of an apoplasmic step at (i) the maternal-filial interface of developing seeds and (ii) the vascular parenchyma-sink cell interface. Circled numbers denote different sink types assigned to each pathway. 1, vegetative apex; 2, elongating axis of a dicotyledonous stem; 3, mature axis of a primary dicotyledonous stem i permanent storage; 4, mature axis of a primary monocotyledonous stem i permanent storage; 5, mature primary root; 6, fleshy fruit; 7, developing seed. ab, apoplasmic barrier; apo, apoplasm; gp, ground parenchyma; sc, sink cell; se—cc, sieve element—companion cell complex; vp, vascular parenchyma.
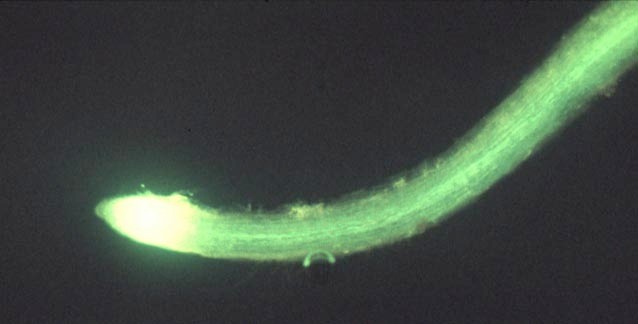
Figure 5.19. Fluorescent micrograph of the distribution of a membrane-impermeant fluorescent dye, carboxyfluorscein (CF), imported through the phloem into roots of French bean (Phaseolus vulgaris L.). The green/yellow fluorescence of CF is confined to the se-cc complexes of mature portions of roots as seen by the thin central band of fluorescence away from the root apex. In contrast, dye spreads through the apex itself apparently via the symplasm of young cells. Scale bar = 2 mm
Photoassimilates can move directly across plasma membranes of se–cc complexes to the surrounding apoplasm (Figure 5.19a). Apoplasmic unloading is important along the axial transport pathway of roots and stems where vascular parenchyma and ground tissues serve as reversible storage sinks.
An entirely symplasmic path of photoassimilate transport from sieve elements to recipient sink cells (Figure 5.18b) operates in a wide range of morphological and metabolic sink types. Terminal growth sinks such as root (Figure 5.19) and shoot apices, as well as vegetative storage sinks such as stems, roots and potato tubers, demonstrate symplasmic unloading.
In most sinks that exhibit symplasmic unloading, photo-assimilates are metabolised into polymeric forms within the recipient sink cells. Sugar cane is a notable exception because it stores sucrose unloaded symplasmically from sieve elements in parenchyma cells of stems. Stem sucrose reaches molar concentrations by this unloading route.
Symplasmic discontinuities exist at interfaces between tissues of differing genomes including biotrophic associations (e.g. mycorrhizas and mistletoes) and developing seeds (Figure 5.18c). In addition, within tissues of the same genome, plasmodesmata can close permanently or reversibly at points along the post-sieve-element pathway. This necessitates photoassimilate exchange between symplasmic and apoplasmic compartments (Figure 5.18c). For instance, photoassimilate exchange between apoplasm and symplasm has been detected in sinks that store high solute concentrations and have unrestricted apoplasmic transport between vascular and storage tissues. Developing seeds, particularly of cereals and large-seeded grain legumes (Patrick and Offler 1995), are another model for symplasmic/apoplasmic pathways.
The apoplasmic space between maternal (seed coat) and filial (embryo plus endosperm) tissues in seeds prevents symplasmic continuity in the unloading pathway (Patrick and Offler 1995). In these organs, photoassimilates are effluxed across membranes of maternal tissues and subsequently taken up across the membranes of filial tissues (Figure 5.18c). Photoassimilates are unloaded from sieve elements and transported symplasmically to effluxing cells where they are released to the seed apoplasm. Influx from the seed apoplasm by the filial generation is restricted to specialised cells located at the maternal–filial interface. The final transport of photoassimilates to the filial storage cells largely follows a symplasmic route.
The symplasm is the most frequently engaged cellular pathway of phloem unloading. Even where an apoplasmic step intervenes (e.g. developing seeds), photoassimilates travel predominantly through the sink symplasm (Figure 5.18c). Symplastic routes do not involve membrane transport and therefore offer lower resistances than apoplasmic routes.
Apoplasmic pathways are restricted to circumstances where (1) symplasmic transport compromises phloem translocation and (2) photoassimilate transport is between genetically distinct (e.g. maternal–filial) tissues. Phloem translocation would be compromised when solutes accumulate to high concentrations in sink cells were it not avoided by symplasmic isolation of phloem from sinks. This is exemplified by the switch to an apoplasmic step during development of tomato fruit. In young fruit, imported sugars are converted into glucose or fructose to support cell division and excess photoassimilate is accumulated as starch. At this stage, phloem unloading of photoassimilates follows a symplasmic route (Figure 5.18b). However, once sugars commence accumulating during cell expansion, apoplasmic transport is engaged (Figure 5.18c). The apoplasmic path isolates pressure-driven phloem import from rising osmotic pressures (P) occurring in fruit storage parenchyma cells (Patrick and Offler 1996).
Radial photoassimilate unloading in mature roots and stems may switch between apoplasmic or symplasmic routes depending upon the prevailing source/sink ratio of the plant. At low source/sink ratios, photoassimilates remobilised from axial stores are loaded into the phloem for transport to growth sinks (Wardlaw 1990). Under these conditions, symplasmic unloading into axial stores might be blocked by plasmodesmal closure while photoassimilates are absorbed by se–cc complexes from the surrounding apoplasm. This would prevent futile unloading while stores are drawn upon. In contrast, net flow of photoassimilates into axial storage pools at high source/sink ratios would be facilitated by plasmodesmal opening.
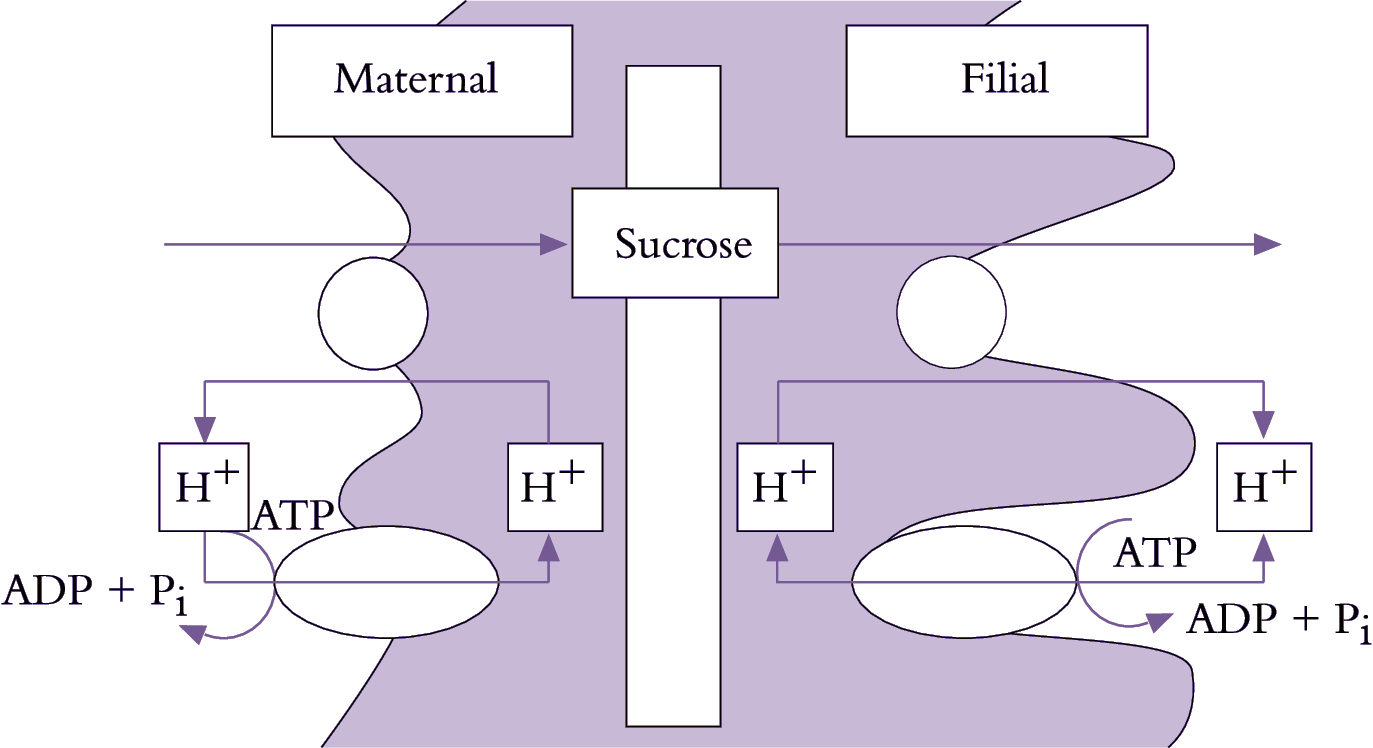
Figure 5.20. Mechanistic model for plasma membrane transport of sucrose from the coat and into the cotyledons of a developing legume seed. Plasma membrane ATPases vectorially pump protons to the seed apoplasm from both the opposing seed coat and cotyledon cells. The proton gradient is coupled to drive sucrose efflux from the seed coats through a sucrose/proton antiporter and sucrose influx into the cotyledons by a sucrose/proton symporter.
Se–cc complexes contain high sugar concentrations (Section 5.2.3(b)). Thus, a considerable transmembrane concentration gradient exists to drive a passive leakage of sugars to phloem apoplasm. Sugars leaked to phloem apoplasm are often retrieved by an active sucrose/proton symport mechanism (Figure 5.20). Thus, net efflux of sugars from se–cc complexes is determined by the balance between a passive leakage and sucrose/proton retrieval.
Passive unloading (Ep) of sucrose from se-cc complexes to the phloem apoplasm (Equation 5.6) is determined by the permeability coefficient (P) of se–cc complex plasma membranes and the transmembrane sucrose concentration (C) gradient between sieve element lumena (se) and surrounding phloem apoplasm (apo).
\[E_p=P(C_{se}-C_{apo}) \tag{5.6} \]
Sinks containing extracellular invertase (e.g. developing tomato fruit, sugar beet tap roots, maize seeds) can hydrolyse sucrose, lowering Capo thereby enhancing sucrose unloading from se–cc complexes. Furthermore, hydrolysis of sucrose renders it unavailable for se–cc complex retrieval by sucrose/proton symport. The resulting hexoses can act as signals to promote cell division in many sinks such as developing seed of Vicia faba.
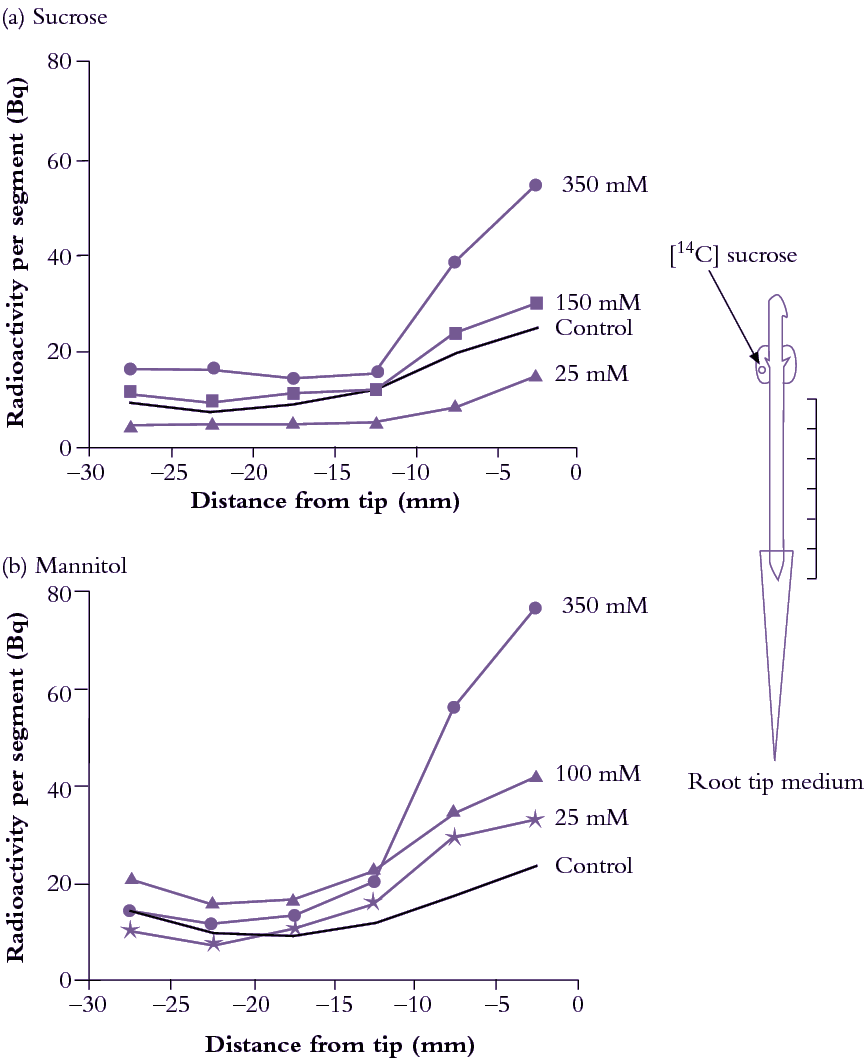
Figure 5.21. Externally supplied solutes have a marked effect on sucrose import into root tips of hydroponically grown pea seedlings. This was tested by immersing root tips in (a) sucrose or (b) mannitol solutions ranging up to 350 mM. Cotyledons, which supply these young roots with carbon, were fed 14C sucrose and 14C arriving in different root parts was measured (Bq per root segment). (a) Import of 14C into root tips was diminished when they were exposed to external sucrose concentrations of less than 100 mM but promoted by sucrose concentrations of 150 to 350 mM; Two effects operate. At low concentrations, sucrose might enter root tip cells and suppress phloem import by a feedback mechanism. At higher concentrations, sucrose might act mainly as an osmoticum (see (b)). Mannitol is not taken up or metabolised quickly and can therefore help answer these questions. (b) Import through the phloem was stimulated by exposing root tips to the slowly permeable sugar mannitol, at concentrations of from 13 to 350 mM. This demonstrates an osmotic dependence of import through the phloem pathway, presumably through progressively decreasing P of root tip cells as external solute concentrations rise (Based on Schultz 1994)
Symplasmic transport is mediated by cytoplasmic streaming in series with intercellular transport via plasmodesmata. Plasmodesmal transport is usually the overriding resistance determining transport rates between cells.
Root tips offer a useful experimental model to explore post-sieve-element symplasmic transport because of morphological simplicity and accessibility. Exposing pea root tips to low sucrose concentrations (<100mM) slowed photoassimilate accumulation (Figure 5.21a) by raising intracellular sucrose concentrations. This response to concentration gradients is consistent with a diffusion component to phloem unloading (Equation 5.7). When roots were bathed in much higher concentrations of either sucrose (Figure 5.22a) or a slowly permeating solute, mannitol (Figure 5.33b), turgor pressure (P) of sieve elements and surrounding tissues decreased and 14C import rose. This is consistent with a hydraulically driven (bulk) flow of photoassimilates into the root apex. Thus, photoassimilate movement from phloem through a symplasmic path can be mediated by diffusion and/or bulk flow. The relative contribution of each transport mechanism depends on the magnitude of concentration and pressure gradients (Equations 5.6 and 5.8).
Physical laws can be used to model diffusion and bulk flow of sucrose through a symplasmic route. Sucrose diffuses through symplasm at a rate (Rd) defined by the product of plasmodesmal number in the path (n), plasmodesmal conductivity to diffusion (Kd) and sucrose concentration difference (ΔC) between sieve elements and sink cell cytoplasm. That is:
\[R_d=n \cdot K_d \cdot \Delta C \tag{5.7} \]
Transport by bulk flow (Rf) is determined by the product of flow speed (S), cross-sectional area of the plasmodesmal flow path (A) and concentration (C) of sucrose transported (Equation 5.2). Flow speed (S), in turn, is a product of hydraulic conductivity (Lp) of a plasmodesma and turgor pressure difference (ΔP) between se–cc complexes and recipient sink cells (Equation 5.8). Flow over the entire pathway considers the number of interconnecting plasmodesmata (n). Thus, bulk flow rate (Rf) is given by:
\[R_f=n \cdot L_p \cdot \Delta P \cdot A \cdot C \tag{5.8} \]
Equations 5.7 and 5.8 predict that sink control of symplasmic photoassimilate transport resides in plasmodesmal conductivity and/or sucrose metabolism/compartmentation.
Sucrose metabolism within sink cells influences cytoplasmic sucrose concentration and Πsink. The difference between Πsink and Πapo determines P (Section 4.3). Sucrose metabolism and compartmentation can affect sucrose concentration gradients and ΔP, both driving forces for symplasmic transport from se–cc complexes to sink cells (Equations 5.6 and 5.8).
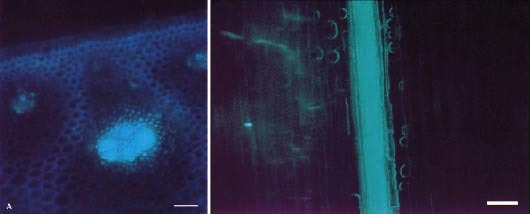
Figure 5.22. Cellular distribution of the apoplasmic tracer fluorescent dye 3-hydroxy—5,8,10—pyren—etrisulphonate (PTS) imported through the xylem in stem explants of sugar cane. (Left) Fluorescent micrograph of a longitudinal section of a stem with PTS (green fluorescence) localised to the vascular bundle. (Right) Fluorescent micrograph of a transverse section showing PTS confined to the vascular bundle. Retention of PTS in vascular bundles demonstrates that a barrier to lateral dye movement must be located in the walls of bundle sheath cells (bs). (Based on Jacobsen et al. 1992)
Transgenic plants which under- or over-express key sugar metabolising enzymes have allowed definitive experiments to be carried out on the role of sucrose metabolism in symplasmic phloem unloading. For example, reduction of sucrose synthase activity (Section 5.4.4) in tubers of transformed potato to 5–30% of wild-type levels depressed dry weight of tubers and starch biosynthesis (Table 5.3). Tubers of transformed plants had very high hexose levels (hence high P) which might contribute to downregulation of photoassimilate import. As a corollary, plants with enhanced starch biosynthesis through overexpression of the key starch synthesising enzyme, ADP-glucose pyrophosphorylase (Section 5.4.5), also had higher rates of photoassimilate import.
For sinks that store sugars to high concentrations (e.g. sugar cane stems), gradients in Π, and hence P, between se–cc complexes and sink storage cells could become too small to sustain transport. Instead, P in the apoplasm of storage tissues increases as sucrose (hence Π) in the storage cell sap rises. This maintains a lower P in storage cells than in sieve elements and sustains transport. High sucrose concentrations in the apoplasm of storage cells is achieved through an apoplasmic barrier which isolates storage parenchyma cells from sieve elements (Figure 5.22).
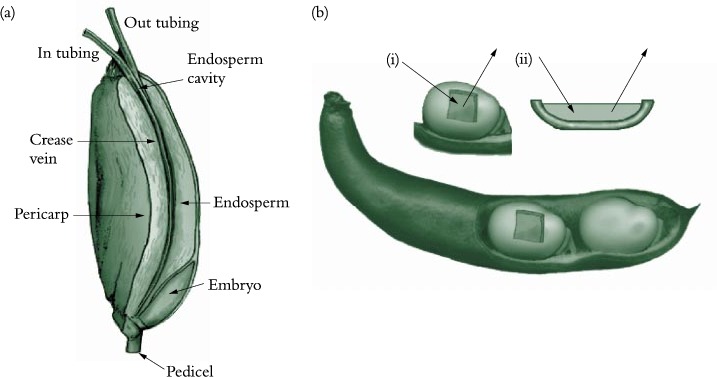
Figure 5.23. Experimental systems used to determine sucrose fluxes in (a) attached caryopses of wheat and (b) coats of developing legume seed. In (a), sucrose effluxed from the maternal tissues was collected by infusing the endosperm cavity of an attached wheat grain with solutions delivered and retrieved through micro-capillaries (Wang and Fisher 1994). In (b), embryos are surgically removed from the coats which may be (i) attached to or (ii) detached from the pod wall. The space vacated by the embryo is filled with a wash solution that is changed at frequent intervals. The wash solution is used to deliver treatments to the seed coat and as a trap to collect the effluxed sucrose.
Phloem unloading in legume seed pods is one case of symplasmic and apoplasmic transport operating in series; the pathway is described in Section 5.4.2(c). Whether sucrose efflux requires energy remains unknown since concentration gradients between seed coats and apoplasm might be steep enough to drive facilitated diffusion. Indeed, using an elegant infusion technique (Figure 5.23a), Wang and Fisher (1994) concluded that efflux from the nucellar projection cells of wheat grain was unlikely to be energy dependent. In contrast, sucrose efflux from coats (maternal tissue) of surgically modified legume seeds (Figure 5.23b) is inhibited by about 50% in the presence of PCMBS, a membrane transport inhibitor. Efflux from legume seed coat cells exhibits charac-teristics of a sucrose/proton antiport. Sucrose uptake by filial tissues is mediated by sucrose/proton symport (Figure 5.23).
A fascinating aspect of phloem unloading in legume seed pods is how photoassimilate demand by filial tissues is integrated with supply from maternal tissues, itself an integration of photoassimilate efflux and import from phloem. One variable that could regulate rates of photoassimilate transport through seed coat symplasm and efflux into apoplasm of the maternal–filial interface is P of seed coat cells (Psc): this would sense depletion of apoplasmic sucrose through uptake by cotyledons, producing a signal in the form of a ΔPsc (Figure 5.24). Specifically, Psc is determined by ΔΠ between the seed coat (Πsc) and seed apoplasm (Πapo), which fluctuates according to photoassimilate withdrawal by cotyledons.
A pressure difference (ΔP) between the points of photo-assimilate arrival (sieve tubes) and efflux (seed coats) drives bulk flow of photoassimilates through the seed coat symplasm. Turgor pressure of seed coat efflux cells is maintained homeostatically at a set point (Pset) by P-dependent efflux into the seed apoplasm. Changes in apoplasmic assimilate concentrations and hence Π are sensed immediately as deviations of Psc from Pset. A rise in Psc produced by photoassimilate depletion around filial tissues elicits an error signal, activating P-dependent solute efflux (Figure 5.25b) and thereby raising photoassimilate concentrations in the apoplasm to meet demand by cotyledons (Figure 5.24c). Long-term increases of sucrose influx by cotyledons, for example over hours, are accompanied by adjustments in Pset (light to dark arrows in Figure 5.24b) which elicit commensurate increases in phloem import rates (light to dark arrows in Figure 5.24a).
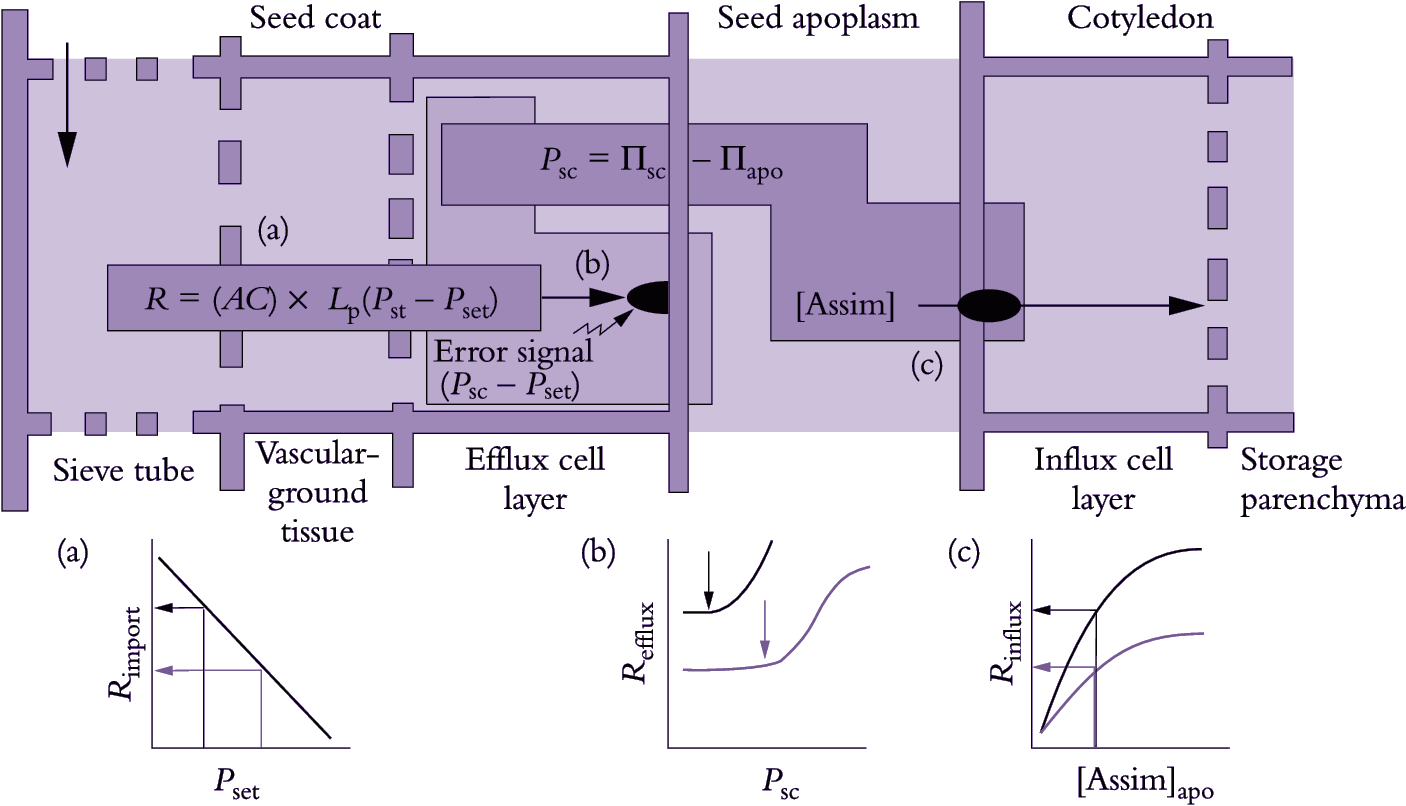
Figure 5.24. A turgor-homeostat model describing the integration of photoassimilate transport to developing legume seeds. Photoassimilate import through phloem (a) and efflux from seed coat to seed apoplasm (b) is mediated by a turgor (P)-dependent efflux mechanism and uptake of sugars by cotyledons (c). Metabolic activity in growing seeds influences sucrose concentrations within cotyledon cells, possibly feeding back on activities of symporters located in plasma membranes of the cotyledon dermal cell complex. Graph (c) denotes increased influx (R) from light to dark curve as sugar demand increases. The apoplasmic solute pool size is small and turns over in less than one hour (Patrick and Offler 1995). Thus, faster photoassimilate withdrawal from the seed apoplasm by cotyledons will rapidly lower apoplasmic osmotic concentration. Since the osmotic difference between seed apoplasm (Πapo) and seed coat (Πsc) is only 0.1-0.2 MPa, a small decrease in osmolality of the apoplasm will elicit a significant increase in seed coat P (Psc).A shift in Psc above the turgor set point (Pset) results in an error signal (see model) which in turn induces an immediate compensatory increase in photoassimilate efflux to the apoplasm (light curve in graph b). Increased photoassimilate efflux acts to maintain a constant Πapo in spite of enhanced flux through the apoplasm (graph c). Consequently, the increased potential for photoassimilate uptake by cotyledons can be fully realised (dark curve in graph c). In the short term (minutes), the turgor-homeostat ensures that Psc is maintained and hence phloem import, which is driven by the turgor difference between sieve tubes and unloading cells (Pst- Psc) is also maintained. Under conditions where cotyledon demand is sustained, Pset in the seed coat adjusts downwards. This results from decreases in Πsc, while Πapo is homeostatically maintained. The decrease in Pset of efflux cells serves to enhance the pressure difference between these cells and the importing sieve elements. As a result, the rate of phloem import into seed coats (Rimport) is increased (graph a, light to dark curve). This new rate of import is commensurate with accelerated sucrose efllux from seed coats to the apoplasm (graph b, light to dark curve) and, ultimately, cotyledons.
The fate of imported photoassimilates depends on sink cell function. In broad terms, imported photoassimilates are primarily used to provide carbon skeletons or signals for growth or storage. Some photoassimilates provide energy for maintenance. Relative flows of photoassimilates to these fates change during cell development and sometimes over shorter time scales depending upon a plant’s physiological state.
Irrespective of sink function, a portion of imported sugars is respired to provide energy (ATP) for maintenance of cell function and structure. Most of this energy is required for continual turnover of cellular constituents such as enzymes and mem-branes. Rates of synthesis and degradation of individual macromolecules vary widely, as does the energy invested in different molecular configurations, so sugar demand for maintenance respiration could differ substantially between tissues.
In growing organs, photoassimilates become substrates for synthesis of new cell material either directly or after biochemical conversions. Other fates for sugars include catabolism in energy-generating pathways which support growth (growth respiration) and storage in vacuolar pools. Stored sugars make an osmotic contribution to growing cells and can act as energy stores in species such as sugar cane. In roots of young barley plants, 40% and 55% of imported sugars are respired and used in structural growth, respectively. Stored sugars turn over each 30 min but account for only 1% of root weight.
In mature cells, imported sugars enter physical (e.g. vacuoles) and chemical (e.g. starch) storage pools with lesser amounts diverted to respiration (15–20%) and structural components. In contrast to growth sinks, stored carbohydrates are ultimately retrieved from storage pools and used by other storage sinks (e.g. germinating seeds) or translocated to support growth and storage processes elsewhere in the plant. Carbohydrate storage can be brief (hours, days) or extend over considerable periods (months to years). Short-term storage of carbohydrates in stems and roots buffers phloem sap sugar concentrations against changes in photoassimilate export from photosynthetic leaves.
Sugars can also be stored in soluble forms by compartmentation into vacuoles. In this case, the tonoplast provides a physical barrier to protect stored sugars from molecular interconversion by cytoplasmic sugar-metabolising enzymes. Vacuolar sugars are accumulated as sucrose, hexoses or fructans (short-chain polymers of fructose). Sucrose and hexoses can accumulate to molar concentrations (0.1–1.5M) in storage parenchyma cells of roots, stems and fruits. For instance, tap roots of sugar beet and stems of sugar cane accumulate 1M sucrose thereby providing 90% of the world’s sucrose. Hexoses are a common form of sugar storage in fruit, contributing to sweetness of edible fruits such as tomato, grape, orange and cucumber. The wine industry depends upon hexoses accumulating to high concentrations (1.5M) in grape berries to fuel fermentation of ‘must’ in wine making. Fructans are stored in significant quantities in leaf sheaths and stems of temperate grasses and cereals. In pasture species, they contribute to forage quality, and in cereals constitute an assimilate pool that is mobilised to support grain filling.
Alternatively, imported sugars may be stored as starch along the axial transport pathway (available for remobilisation to buffer phloem sap sugar concentrations) or in more long term storage pools of terminal sink organs such as tubers, fruits and seeds. The proportion of photoassimilates diverted into starch differs widely between species and organs. Starch accounts for some 90% of dry weight of potato tubers and cereal grains.
The chemistry of storage products can change during organ development. For instance, starch is the principal storage carbohydrate in young tomato fruit. Later in fruit development, stored starch is hydrolysed and contributes to hexose accumulation in vacuoles of fruit storage parenchyma cells. In other fruits, significant switches between hexose and sucrose accumulation occur during development. All these changes are brought about by ontogenetic shifts in activities of sugar-metabolising enzymes.
Phloem-imported sucrose can reach the cytoplasm of recipient sink cells chemically unaltered or be hydrolysed en route by extracellular invertase into its hexose moieties. These sugars may then enter a number of metabolic pathways or be compartmented to vacuolar storage (Figure 5.25).
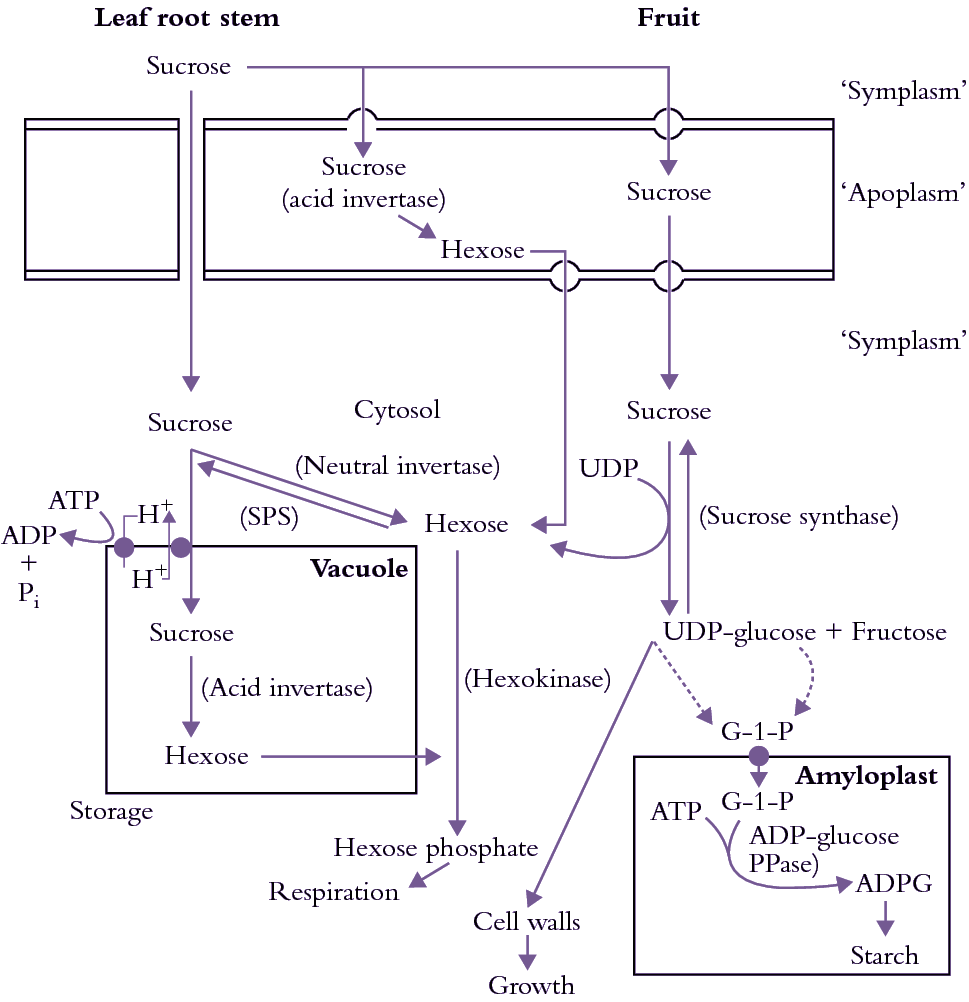
Figure 5.25. Pathways of sugar metabolism and compartmentation within sink cells. Sugars can be delivered to sink cells through either apoplasmic or symplasmic pathways. Within the sink apoplasm, sucrose can be hydrolysed to hexoses by an extracellular invertase. Apoplasmic sugars are transported across plasma membranes of sink cells by proton/ sugar symporters. Alternatively, sucrose enters the sink cytoplasm through a symplasmic path. Within the sink cytoplasm, sucrose can be hydrolysed or compartmented into vacuolar storage. Sucrose hydrolysis provides substrates for energy metabolism or for synthesis of macro-molecules. Invertase activity is important to sustain hexose supply for glycolysis. Sucrose synthesis from the hexose pool is catalysed by sucrose phosphate synthase (SPS). Degradation of sucrose by sucrose synthase generates fructose and uridine diphosphate glucose (UDP-glucose) which enters various biosynthetic pathways including cellulose and starch synthesis. In the case of starch biosynthesis, UDP-glucose and fructose generate glucose-1-phosphate (G-1-P) which is transported across the amyloplast membrane. Accumulated G-1-P is interconverted into adenine diphosphate glucose (ADP-glucose) by the enzyme adenine diphosphate glucose pyrophosphorylase (ADP-glucose PPase). ADP-glucose is the substrate for starch polymer formation. Sucrose compartmentation into the vacuole is mediated by a sucrose/ proton antiporter located on the tonoplast. Within the vacuole, sucrose can be exposed to invertase hydrolysis with the hexose products accumulating or leaking back to the cytoplasm
Sucrose is metabolically inert and, in order to be metabolised, must be hydrolysed to glucose and fructose. Only two enzymes are capable of metabolising sucrose in green plants. These are invertase and sucrose synthase (Figure 5.25) and they are paramount in sugar metabolism after phloem unloading.
Invertase catalyses irreversible hydrolysis of sucrose to its hexose moieties, glucose and fructose. Both acid and neutral invertases occur in plants, with pH optima of about 5 and 7.5, respectively. The activity of invertases varies with plant species, organ type and stage of development. Acid invertases, located in cell wall or in vacuole, are usually active in rapidly growing leaves, stems and fruits and seeds (Ruan et al. 2010), making hexoses available for regulating gene expression and for respiration and biosynthesis. Reduced acid invertase activity in vacuoles during development of sugar cane stems, and its absence from sucrose-accumulating tomato fruit, is a major factor in sucrose accumulation in vacuoles of these tissues. Suppression of cell wall invertase activity led to shrunken seed in maize and small fruit in tomato and loss of pollen fertility in tomato, wheat and rice, demonstrating its critical roles in these reproductive organs. Less is known about the physiological role of neutral invertases..
Sucrose synthase is mainly located in the cytoplasm but recent research also shows that the enzyme may also be associated with plasma membrane and even present in cell wall matrix. It catalyses sucrose cleavage to fructose and UDP-glucose, a high-energy ester of glucose. UDP-glucose is a substrate for biosynthesis of cellulose and may be converted further for starch synthesis.High activities of sucrose synthase are found in both growing and starch storage tissues. In the cytoplasm of starchy tissues, UDP-glucose is converted by UDP-glucose pyro-phosphorylase to glucose-1-phosphate, which is transported across amyloplast membranes. In amyloplasts, glucose-1-phosphate provides glucose moieties for starch synthesis in a pathway comparable to starch formation in chloroplasts of photosynthetic leaves. The critical role of sucrose synthase in starch synthesis is demonstrated with potatoes transformed with an antisense construct of the gene encoding tuber-specific sucrose synthase. Tuber sucrose synthase activity in transformed plants was depressed significantly while the activities of key starch biosynthetic enzymes were unaltered. Low sucrose synthase activity was directly responsible for a proportional decrease in starch accumulation (Zrenner et al. 1995).
Sinks that accumulate soluble sugars have predictably low sucrose synthase activities. Contrastingly high sucrose synthase activities in phloem vessels may be responsible for energy production for phloem loading or unloading and maintaining cellular function of companion cells.
Hexoses transported to the sink cytoplasm are rapidly phosphorylated to hexose-6-phosphates by glucose- and fructose kinases. In these forms, hexoses can be used as substrates for respiration or for synthesis of new cell constituents. Alternatively, sucrose phosphate synthetase can convert them to sucrose, as in leaves (Chapters 1 and 2). Sucrose synthesized by this reaction can be accumulated in vacuoles (e.g. sugar beet tap roots, sugar cane stems) or be rehydrolysed into hexoses by a vacuolar acid invertase (e.g. grape berries).
The pressure-flow hypothesis provides a compelling model to explain sink strength in plants. Evidence such as accelerated import of photoassimilate into roots with artificially lowered P lends empirical support to the model.
Knowledge of cellular and molecular events in phloem unloading and photoassimilate use begins to reveal the array of control steps which underlie photoassimilate unloading and relative sink strength. Photoassimilate import into sinks by apoplasmic pathways or by diffusion through symplasmic pathways (Figure 5.19) is controlled by P in se–cc complexes. In contrast, when phloem unloading is by bulk flow through a symplasmic route (e.g. legume seed coats), P in cells responsible for photoassimilate efflux to the apoplasm controls unloading. Unloading into storage tissues is controlled by P in sink (storage) cells.
These processes at the cell and tissue level must now be related to a whole-plant perspective of sink control of photoassimilate partitioning, taking into account influences of plant development and environmental factors. How plants use photoassimilates (e.g. switch from growth to storage) is accompanied by alterations in the cellular pathway of import. Phytohormones also play a role in photoassimilate partitioning through their influence on development and intercellular signalling.
Plant development generates new sinks, for example in meristems where cells undergo division or growth zones where enlarging cells import photoassimilates.
Potential sink size is set largely during the meristematic phase of development through determination of total cell number per organ. Photoassimilate supply has been implicated as a limiting factor in initiation of leaf primordia at the apical dome and subsequent early development by cell division. Substrate supply for developing seeds (endosperm, embryo) and root and floral apices might also be restricted.
Agricultural yields might therefore increase if plants could be modified to enhance the supply of photoassimilates to meristems. Which factors regulate photoassimilate supply to meristematic sinks? Rate equations describing mass flow of phloem sap (Section 5.2.5) predict that photoassimilate supply reaching a sink will be determined by source output (setting photoassimilate concentration in sap and P in sieve elements at the source) and modulated by Lp of the transport pathway. Increased photoassimilate output from source leaves increases growth activity of primary meristems. Even during reduced source output, photoassimilate import and meristematic sink strength can be maintained by remobilisation of storage reserves. Manipulating competition for photo-assimilates by more established sinks also suggests that source output influences sink behaviour.
Cultivated plants demonstrate these principles. For example, flushing CO2 into glasshouses increases flower set and hence yield of floral and fruit crops. Similarly, applying growth regulators to induce abscission of some floral apices lessens the number of sinks competing for photoassimilates at fruit set and leads to larger and more uniform fruit at harvest. Alternatively, breeding programs have reduced sink strength of non-harvestable portions of crops and hence the severity of competition. For instance, breeding dwarf varieties of cereals has reduced photoassimilate demand by stems with a consequent increase in floret numbers set and grain size.
These observations imply that increases in net leaf photosynthesis and phloem loading should set higher yield potentials. Yet meristems import only a small proportion of total plant photoassimilate. It may be that phloem conductance limits photoassimilate delivery to meristems; increases in source output would amplify the driving force for transport and hence bulk flow through a low-conductance pathway.
Given that mature phloem pathways have spare transport capacity (Section 5.2.5), any transport limitation imposed by low path conductance might be expected within immature sinks. Photoassimilate import into meristematic sinks involves transport through partially differentiated provascular strands that might extend up to 400µm. Movement through this partially differentiated path is symplasmic (Section 5.2.2(b)). Hence, plasmodesmal numbers and transport properties of plasmodesmata could play a critical role in photoassimilate supplies to sinks and determination of sink size (Equation 5.7).
As cells expand and approach cell maturity, photoassimilates are increasingly diverted into storage products. Towards maturity, fully differentiated phloem pathways with spare transport capacity link expansion/storage sinks with photosynthetic leaves. Photoassimilate import by these sinks depends on duration of the storage phase. This can be short term for sinks located along the axial transport pathway and long term for sinks sited at the ends of transport pathways (e.g. tubers, fruits and seeds).
Storage along the axial pathway occurs mainly when photoassimilate production exceeds photoassimilate demands by terminal sinks. However, storage is not necessarily a passive response to excess photoassimilate supply. Stems of sugar cane store large quantities of photoassimilates (50% of dry weight is sucrose) even during rapid growth of terminal sinks. Photoassimilates might be stored as simple sugars (e.g. sugar cane stems) or as polymers (fructans in stems of temperate grasses; starch in stems and roots of subtropical cereals, herbaceous annuals and woody perennials). Photoassimilates stored along axial pathways buffer against diurnal and more long term fluctuations in photoassimilate supply to terminal sinks. In woody deciduous species, axially stored photoassimilates also provide a long-term seasonal storage pool that is drawn on to support bud growth following budburst. Remobilised photoassimilates can contribute substantially to biomass gain of terminal sinks. For instance, in some mature trees, over half the photoassimilates for new growth come from remobilised reserves; similar proportions of stem-stored fructans contribute to grain growth in cereals when photoassimilate production is reduced (e.g. by drought). Physiological switching between net storage and remobilisation is an intriguing regulatory question.
Growth and development of meristems is determined by phloem unloading events and metabolic interconversion of photoassimilates within recipient sink cells. These transport and transfer processes vary between sinks and can alter during sink development. Techniques now exist to alter expression of membrane porter proteins and possibly enhance photoassimilate import by sinks such as seeds which have an apoplasmic step in the phloem unloading pathway. Prospects of altering plasmodesmal conductivity will improve once plasmodesmal proteins are identified and their encoding genes known.
Phloem unloading of nutrients follows water release from vascular system.
As discussed before, because of their low transpiration rates, developing sinks typically import water through phloem, not xylem. Water unloaded from phloem is used for cell growth or recycled back to the parental bodies. Water is unloaded from se-cc complexes symplasmically in majority of sinks by bulk flow. For growth sinks such as shoot or root apices, continued symplasmic flow of phloem-imported water can drive cells expansion. In post-phloem unloading pathways interrupted with an apoplasmic step, water must exit cells of the unloading path across cell membranes, facilitated by aquaporins (AQPs). AQPs responsible for water flow across cell membranes are plasma membrane intrinsic proteins (PIPs) and tonoplast intrinsic proteins (TIPs). Strong PIP expression in expanding post-veraison grape berries has been shown to correlate with water flows into, and from, the berry apoplasm. For sinks that stop expansion but continuously accumulate biomass, water transported to storage sink apoplasms is recycled back to the parent plant body through a xylem route. Important roles played by AQPs in water recycling are indicated by their high expression at this stage in developing seed, particularly in the vascular parenchyma cells.
In conclusion, this chapter has shown how growth and development of meristems and other sinks is determined by phloem unloading events, and metabolism of the assimilates within the recipient sink cells. These transport and transfer processes vary between specific sinks, and can alter during development. Molecular techniques that alter expression of membrane transporters can be used to study the pathways and limitations of photoassimilate transport into sinks such as seeds that have an apoplasmic step in the phloem unloading pathway, with the possibility of enhancing the rate of grain growth and crop yield in the future.
Aoki K, Suzui N, Fujimaki S et al. (2005) Destination-selective long-distance movement of phloem proteins. Plant Cell 17: 1801-1814.
Brenner M (1987) The role of hormones in photosynthate partitioning and seed filling. In PJ Davies, ed, Plant Hormones and their Role in Plant Growth and Development, Kluwer, Dordrecht, pp 474-493
Canny MJ (1973) Phloem Translocation. Cambridge University Press
Chen LQ, Qu XQ, Hou BH, Sosso D, Osorio S, Fernie AR, Frommer WB (2012) Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 335: 207-211
Covarrubias AA, Reyes JL (2010) Post-transcriptional gene regulation of salinity and drought responses by plant microRNAs. Plant Cell Environ 33:481-489.
Daie J, Watts M, Aloni B, Wyse RE (1986) In vitro and in vivo modification of sugar transport and translocation in celery by phytohormones. Plant Sci 46: 35-41
Fisher DB, Frame JM (1984) A guide to the use of the exuding stylet technique in phloem physiology. Planta 161: 385–393
Frommer WB, Hirner B, Kuhn C, Harma K, Martin T, Reismeier JW, Schulz B (1996) Sugar transport in higher plants. In M Smallwood, JP Knox, DJ Bowles, eds, Membranes: Specialised Functions in Plants. Bioscientific, Oxford, pp 319-335
Geiger DR (1975) Phloem loading. In Zimmerman MH, Milburn JA, eds, Transport in Plants 1. Phloem Transport. Springer-Verlag, Heidelberg, pp 395-431
Grusak MA, Beebe DU, Turgeon R (1996) Phloem loading. In Zamski E, Schaffer AA eds, Photoassimilate Distribution in Plants and Crops. Source-Sink Relationships. Marcel Dekker, New York, pp 209-228
Jacobsen KR, Fisher DG, Maretzki A, Moore PH (1992) Developmental changes in the anatomy of the sugarcane stem in relation to phloem unloading and sucrose storage. Bot. Acta 105: 70-80
Kallarackal J, Milburn JA (1984) Specific mass transfer and sink-controlled phloem translocation in castor bean. Aust J Plant Physiol 11: 483-490
Kriedemann P, Beevers H (1967) Sugar uptake and translocation in the castor bean seedling. II. Sugar transformations during uptake. Plant Physiol 42: 174-180
Lough TJ, Lucas W. (2006) Integrative plant biology: role of phloem long-distance macromolecular trafficking. Annu Rev Plant Biol 57: 203–232
Milburn JA, Baker DA (1989) Physico-chemical aspects of phloem sap. In DA Baker, JA Milburn, eds, Transport of Assimilates. Longman, Harlow, pp 345-359
Milburn JA, Kallarackal J (1989) Physiological aspects of phloem translocation. In DA Baker, JA Milburn, eds, Transport of Assimilates. Longman, Harlow, pp 264-305
Moorby J, Jarman PD (1975) The use of compartmental analysis in the study of movement of carbon through leaves. Planta 122: 155-168
Münch E (1930) Die Stoffbewegungen in der Pflanze. Fischer, Jena
Pate JS, Peoples MB, Atkins CA (1984) Spontaneous phloem bleeding from cryopunctured fruits of a ureide-producing legume. Plant Physiol 74: 499-505
Patrick JW (1997) Phloem unloading: sieve element unloading and post-sieve element transport. Annu Rev Plant Physiol Plant Mol Biol 48: 191–222
Patrick JW, Offler CE (1995) Post-sieve element transport of sucrose in developing seed. Aust J Plant Physiol 22: 761-769
Patrick JW, Offler CE (1996) Post-sieve element transport of photoassimilates in sink regions. J Exp Bot 28: 736-743
Rodriguez-Medina C, Atkins CA, Mann A, Jordan M, Smith PMC (2011) Macromolecular composition of phloem exudate from white lupin (Lupinus albus L.). BMC Plant Biol 11: 36-54
Ruan YL, Jin Y, Yang YJ, Li GJ, Boyer JS (2010) Sugar input, metabolism, and signaling mediated by invertase: roles in development, yield potential, and response to drought and heat. Mol Plant 3: 942-955
Schulz A (1994) Phloem transport and differential unloading in pea seedlings after source and sink manipulations. Planta 192: 239-249
Turgeon R (2010) The role of phloem loading reconsidered. Plant Physiol 152: 1817-1823
van Bel AJE (1993) Strategies of phloem loading. Annu Rev Plant Physiol Plant Mol Biol 44: 253-282
Wardlaw IF (1990) The control of carbon partitioning in plants. New Phytol 27: 341-381
Wang N, Fisher DB (1994) Monitoring phloem unloading and post-phloem transport by microperfusion of attached wheat grains. Plant Physiol 104: 7-16
Wright JP, Fisher DB (1980) Direct measurement of sieve tube turgor pressure using severed aphid stylets. Plant Physiol 65: 1133-1135
Varkonyi-Gasic E, Gould N, Sandanayaka M, Sutherland P, MacDiarmid RM (2010) Characterisation of microRNAs from apple (Malus domestica ’Royal Gala’) vascular tissue and phloem sap. BMC Plant Biol 10: 159-173
Zrenna R, Salanoubat M, Willmitzer L, Sonnewald U (1995) Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L). Plant J 7: 97-107


