Chapter editor: Dennis H. Greer
School of Agricultural and Wine Sciences, Charles Sturt University, Australia
This Chapter is updated from the original chapter (1st edition) from Sharon Robinson, Jenny Watling, Dennis Bittisnich, Shu Fukai, Chris Beadle, Mike Clearwater and Paul Kriedemann.
12.0-Ch-Fig-12.0-NoNum.png

Shafts of sunlight penetrate a natural forest of Norfolk Island pine (Araucaria heterophylla). (Photograph courtesy D.H. Ashton)
Terrestrial ecosystems are both sustained and regulated by sunlight: sustained in massive ways by photosynthetically active radiation, but regulated in subtle ways by other wavelengths. Wavelengths most effective for photosynthesis occupy a band between about 380 and 720 nm. A wider band from about 350 to 800 nm spans the action spectra for other crucial responses in plant growth and reproductive development that are also light regulated. These include seed germination, tropisms, morphogenesis, pigmentation, and photoperiodic responses such as floral initiation (topics covered in Chapter 8 - environmental effects on plant development).
The outcomes of these two light-dependent categories differ by many orders of magnitude in terms of energy flow within a plant community. In one case, a flow of radiant energy is converted into chemical energy and stored as biomass; in the other, miniscule levels of radiant energy trigger shifts in gene expression and consequent developmental responses. Nevertheless, each category is mediated by pigment systems that transduce solar energy into highly ordered chemical forms: biosynthetic systems in photosynthesis, triggering systems in photo-morphogenesis (light-mediated development, where plant growth patterns respond to the light spectrum).
12.0-Ch-Fig-12.1.png
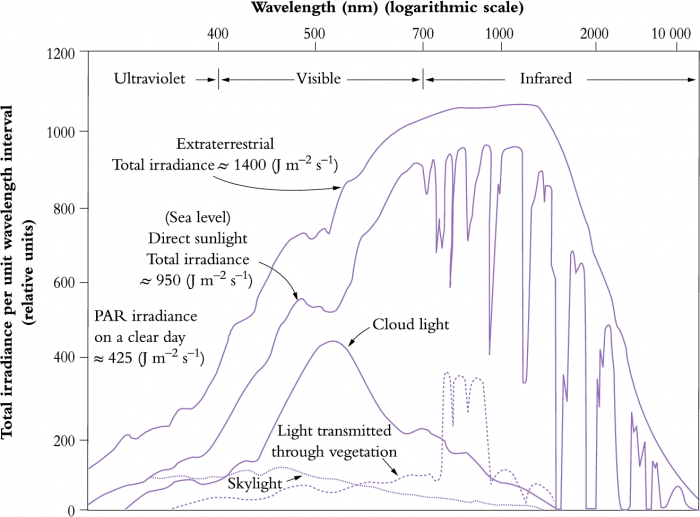
Figure 12.1 Sunlight passing through the earth's atmosphere is altered both in quantity and spectral composition.
The earth’s atmosphere attenuates solar radiation in highly selective ways (Figure 12.1). Substantial amounts of infrared energy (between about 850 and 1300 nm) are absorbed by CO2, ozone and especially by water vapour, while ozone is principally responsible for a cut-off in ultraviolet radiation below about 300 nm. Attenuation of discrete bands of infrared radiation by water vapour is especially noticeable (right side of figure), while absorption of ultraviolet by stratospheric ozone is also of special significance for terrestrial organisms. Cloud light is especially rich in visible wavelengths with a peak around 500 µm. Our atmosphere thus represents a ‘window’ through which visible wavelengths pour onto the earth’s surface, and terrestrial life forms have evolved with attributes that are a direct consequence of this spectral composition.
Selective filtering of wavelengths either side of the visible spectrum is crucial. Ultraviolet radiation is comprehensively absorbed by biological ingredients, especially proteins, RNA and DNA and pigments. Because energy per quantum increases with the decrease in wavelength, ultraviolet radiation imposes a heavy load of energy on biological components with attendant disruption. Similarly, wavelengths beyond the visible spectrum, though less energetic, are still damaging because tissue water absorbs infrared radiation. All life processes operate within an aqueous milieu, so that the absorption properties of water molecules would put cellular function at risk if sunlight was not also attenuated with respect to infrared radiation by the atmosphere.
Between these two extremes stands the visible spectrum, and it is no coincidence that all manner of biological systems have evolved to make effective use of this narrow band of solar radiation. Vegetation shows strong attenuation of visible wavelengths that drive photosynthesis, but transmit near-infrared radiation that would otherwise heat leaves. Vascular plants are a case in point, where canopy, leaf and chloroplasts have all adapted to the light climate with features that optimise their use of sunlight. These include mechanisms to deal with excess radiation.
Photosynthetic efficiency in low light confers a selective advantage on shade-adapted plants, but also renders them especially vulnerable to full sun. Accordingly, such species have evolved remarkable features for photoprotection. Their acclimation to sun and shade, together with properties of sun-loving plants, thus reveal an extraordinary plasticity in the photosynthetic apparatus of vascular plants (Section 12.1). Even increased UV-B radiation, commonly associated with global change, and the so called ‘ozone hole’ over Antarctica, elicits responses that offer photoprotection to plants (Section 12.6).
Aside from the biological hazards, solar radiation obviously sustains global photosynthesis, and close quantitative relationships exist between energy absorption and biomass production. Such relationships are especially well defined for managed communities, with cropping, plantation forests and horticulture providing clear examples (Sections 12.2, 12.3, 12.4). Canopy architecture is a major determinant of sunlight interception, and hence production of biomass. Canopy pruning in horticulture are examples of enhanced productivity through increased interception of light. In such cases, light-dependent regulation of plant development assumes prominence because tree and vine canopies are shaped for both overall interception of light as well as maximum fruitfulness.
A note on units: Visible wavelengths of sunlight can be represented as either a quantum flux or a radiant energy flux. Quantum flux is regarded here as synonymous with 'photon irradiance' (Q) and has units of µmol quanta m-2 s-1 ('µmol quanta' rather than 'µmol photons' because the quantum energy derived from photons drives photosynthesis). For the sake of making a clear distinction from quantum flux, radiant energy flux is simplified to 'irradiance', and for present purposes, irradiance coincides with photosynthetically active radiation (PAR). Irradiance is then expressed as joules (J) per square meter per unit time. Depending on the application, time can span seconds, days or years, and is then coupled with either joules, megajoules (MJ) or gigajoules (GJ).