CO2 fixed by photosynthesis in chloroplasts has several possible fates, but most ends up as sucrose or starch. Starch is stored in chloroplasts, and sucrose is stored in vacuoles of mesophyll cells. Both starch and vacuolar sucrose serve as temporary storage pools from which the cytoplasmic sucrose pool is replenished. Sucrose, along with amino acids and mineral nutrients, is loaded into the phloem tissue which consists of sieve element—companion cell (se—cc) complexes for long-distance transport to growing tissues and other non-photosynthetic sinks. These solutes are exchanged reversibly between se-cc complexes and short- and long-term storage pools along the axial pathway. Short-term storage pools include phloem apoplasm, and the protoplasm of non-transport cells provides a long-term storage pool. At the end of the pathway, sucrose and other transported solutes are consumed in respiration and growth, or are stored as solutes in vacuoles or polymers in amyloplasts (starch) or protein bodies.
The overall flow of photoassimilates throughout the plant can therefore be called a source–path–sink system (Figure 5.1).
Fig5.13-new-p.png
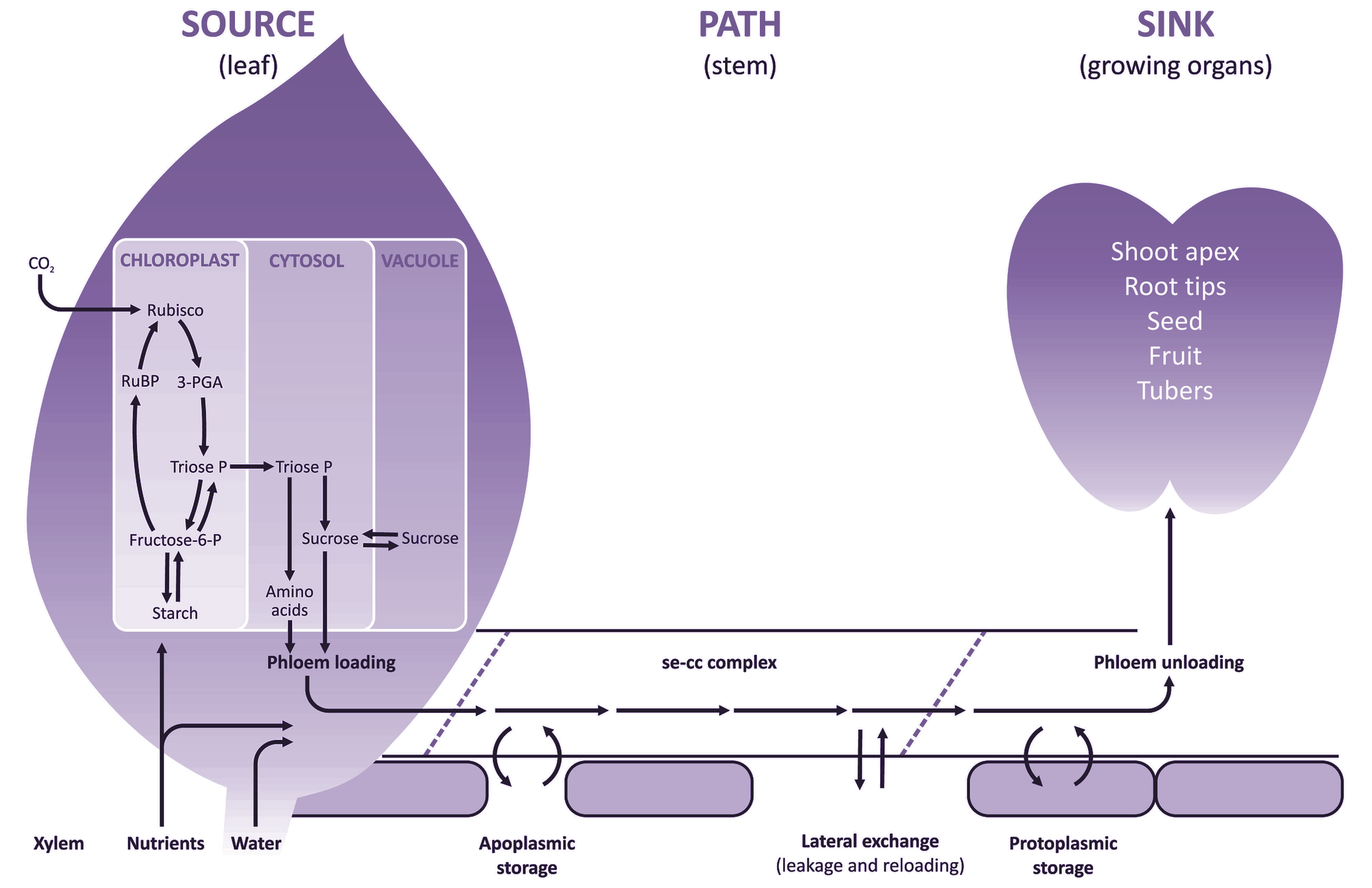
Figure 5.1. Schematic diagram of transfer and transport processes contributing to the flow of assimilates acquired from aerial or soil environments, through the source-path-sink system. CO2 fixed by photosynthesis in chloroplasts gives rise to sucrose and starch. Sucrose, amino acids and mineral nutrients are loaded into sieve element—companion cell (se—cc) complexes of leaf phloem for long-distance transport to non-photosynthetic sinks. These solutes are exchanged reversibly between se-cc complexes and short- and long-term storage pools along the axial pathway. Short-term storage pools include phloem apoplasm, whereas the protoplasm of non-transport cells provides a long-term storage pool. In sink tissues, solutes are used for respiration, growth or storage.