Formation of gaps provides an important opportunity for many rainforest plants to escape from the dim understorey environment and reach maturity as canopy trees. Rainforest habitats actually present a continuum of light availability ranging from almost total shade (diffuse light) through intermediate levels of direct and diffuse sunlight to a wide gap where direct sunlight is received for most of a day. Input of direct sunlight beneath a closed canopy can be surprisingly high because of sun patches and more transient sunflecks. Sun patches occur when small and variable openings in the overlying canopy permit direct sunlight to penetrate to the forest floor, resulting in the familiar patchwork of sunlight and shade which can be seen in any understorey on a clear day (Figure 12.16).
12.1-Ch-Fig-12.16.png

Figure 12.16 Cohort of Argyrodendron sp. seedlings in a protected niche at the base of a mature tree of that same species growing in a rainforest on Atherton Tableland. Scale bar = 15 cm. (Photograph courtesy P.E. Kriedemann)
Competition for light is intense, and a sunfleck shown passing over this tiny population would be providing a much-needed source of sunlight, namely a few minutes of around 1,500 µmol quanta m-2 s-1 compared with background photon irradiance of around 150 µmol quanta m-2 s-1.
Patches of sunlight move across the forest floor on bright days, and will illuminate any leaves or parts of leaves which lie in their path (Figure 12.17). Daily total photon irradiance on the occasion of the measurement shown in Figure 12.17 was 6.0 mol quanta m-2 d-1). The abrupt increase from a background level of around 50 µmol quanta m-2 s-1 to 1,750 µmol quanta m-2 s-1 energises photosynthesis, but is counteracted by increased leaf temperature, especially during prolonged exposures. Transpiration cooling is a significant component of heat budgets for such large leaves, so that adequate soil moisture is a prerequisite for continuing leaf gas exchange during a sunfleck.
12.1-Ch-Fig-12.17.png
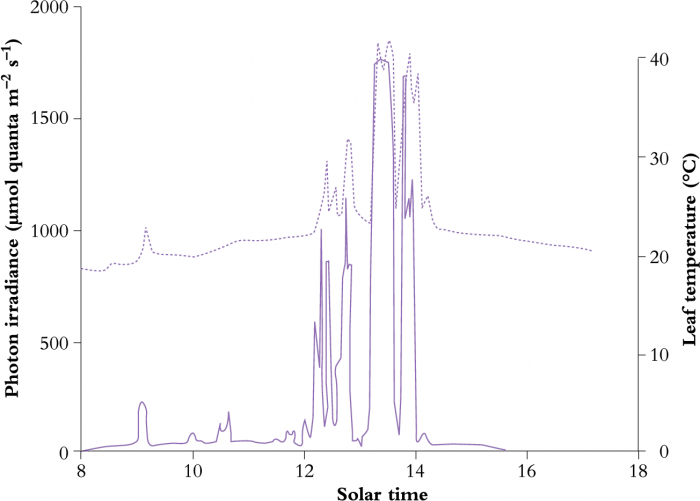
Figure 12.17 Natural variation in photon irradiance and leaf temperature experienced by Alocasia macrorrhiza growing in the understorey of a North Queensland rainforest. (J.R. Watling et al. 1997a, b)
Sunfleck frequency will be an additional factor for carbon gain during exposure to strong photon irradiance. Photosynthetic induction state diminishes to a minimum after 30 min in low light (see Figure 12.18 below) and some minutes are required to regain full capacity in strong light. Infrequent sunflecks are thus used with reduced efficiency.
Sunflecks and sun patches are of potential use to understorey plants for photosynthesis, but is this potential realised? Growth of understorey tree seedlings has been shown to be correlated with the amount of direct light received in sunflecks, and up to 60% of carbon gain in such environments has been attributed to this source. However, when compared with expected values based on the known steady-state response of plants to light, sunfleck utilisation is often below predicted values (Pfitsch and Pearcy 1989). Moreover, species vary in their capacity to utilise sunflecks. Watling et al. (1997a) measured the growth of four Australian rainforest species under simulated sunfleck regimes and showed that sunflecks contributed to growth in two species (Diploglottis diphyllostegia and Micromelum minutum), whereas the other two species (Alocasia macrorrhiza and Omalanthus novo-guinensis) were unable to make effective use of sunflecks.
Factors underlying this variation in sunfleck utilisation efficiency
Two components of a plant’s photosynthetic physiology will determine how the light in a sunfleck is used. Firstly, photosynthetic capacity (Pmax) will set a ceiling on the amount of light a plant can use. Secondly, a few minutes of illumination at least are needed for PCR cycle intermediates to reach critical levels, and this ‘induction requirement’ of photosynthesis determines how quickly a leaf can respond to an increase in photon irradiance.
Measured photosynthetic capacities of understorey plants are often low. When a leaf experiences a sunfleck, carbon fixation will increase to the point of light saturation. If an understorey plant could increase Pmax, it could utilise more light. But there are trade-offs. For example, higher respiration rates would increase the light-compensation point, and increase carbon losses during the low light periods separating sunflecks.
Regardless of photosynthetic capacity, the ability of a plant to use sunflecks is also affected by the induction requirement of photosynthesis. When a leaf that has been in low light for some time is exposed to an increase in photon irradiance, the rate of photosynthesis does not increase instantaneously to the new level. Instead, there is a gradual increase in assimilation which can take from 10–60 min for completion. This ‘induction period’ varies according to species as well as the induction state of the leaf concerned. Three different processes are involved; namely (1) buildup of PCR cycle intermediates and in particular ribulose-1,5-bisphosphate (RuBP), (2) light-dependent activation of Rubisco, and (3) light-dependent opening of stomata. Each of these processes follows a different time-course. Buildup of a metabolite pool is fastest (1–2 min), followed by Rubisco activation (2–5min) and finally stomatal opening (up to 60 min). Relaxation in low light is more protracted but generally occurs in the same sequence, leading to a decline in the induction state.
12.1-Ch-Fig-12.18.png
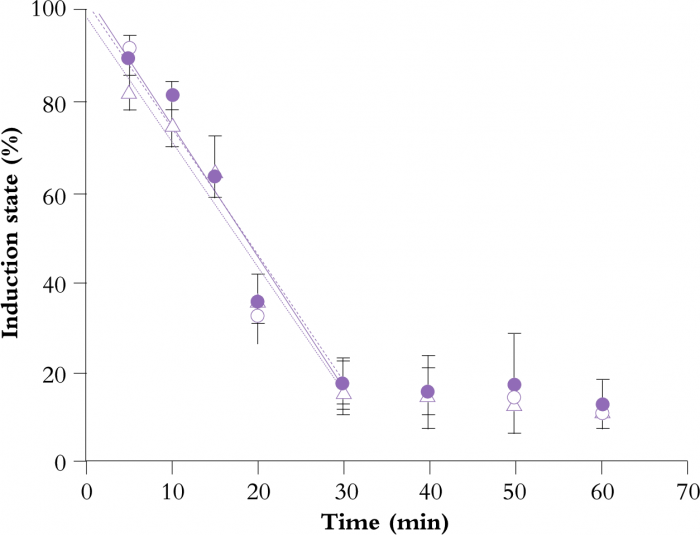
Figure 12.18 Loss of photosynthetic induction state in Alocasia macrorrhiza following transfer from saturating light to low light. Replicate determinations represented by different symbols and error bars. (J.R. Watling et al. 1997a, b)
Thus, the longer a leaf has been in low light, the lower it’s the induction state. Figure 12.18 illustrates this for a leaf of Alocasia macrorrhiza, a plant common in rainforest understoreys and forest edges in eastern Australia. Fully-induced leaves were transferred to low light (10 µmol quanta m-2 s-1) for different lengths of time (up to 60 min). Their induction state was determined as the proportion of light-saturated photosynthetic capacity (Pmax) achieved within 2 min after return to saturating light. Induction loss in Alocasia, therefore, has a half time of about 25 min, but other species have been found to be either faster (e.g. Adenocaulon bicolor, an understorey herb from the redwood forests of western USA; Pfitsch and Pearcy 1989) or slower (e.g. Castanospora alphandii, a shade-tolerant tree from Australian rainforests; Watling et al. 1997a, b).
Chazdon and Pearcy (1986) showed that continuous light is not needed for induction to proceed. If leaves were subjected to a series of 60 s lightflecks (artificial sunflecks) separated by 2 min of low light, then induction state increased with each successive lightfleck. In nature, sunflecks are often clustered with a sequence of irregular bursts, separated by dull periods of variable duration. Under these circumstances, the state of induction might improve such that a plant will respond more rapidly to closing sunflecks in each sequence. This is supported by the data of Chazdon and Pearcy (1986), where the efficiency with which a sequence of lightflecks was utilised increased with successive lightflecks (efficiency was calculated as the actual amount of carbon fixed during a lightfleck relative to the amount predicted if there had been no induction period).
Carbon gain by fully induced leaves during lightflecks can exceed that expected, resulting in improved efficiency of light utilisation. This ‘intermittency phenomenon’ was noted by Kriedemann et al. (1973) in grapevine leaves exposed to high-frequency lightflecks. Similar investigations in a number of other species have also shown that such enhancement occurs only with short-duration lightflecks and is more prominent in fully induced leaves. Alocasia macrorrhiza showed improved efficiency of lightfleck utilisation by fully induced leaves only when lightflecks were of 40 s duration or less. Un-induced leaves needed lightflecks 10s or shorter (Chazdon and Pearcy 1986).
This apparent improvement in light use efficiency results from continued carbon fixation in low light (or darkness) following a lightfleck. During a short lightfleck, pools of triose phosphate and RuBP build up because carbon fixation runs transiently slower than the light reactions. This pool of PCR intermediates is then used for post-illumination CO2 fixation. In rainforest understoreys, where sunflecks are generally longer than a few seconds, this kind of enhancement is unlikely to be important. However, it may contribute significantly to carbon gain under crop canopies, where sunflecks are much shorter and more frequent.
One consequence of generally low photosynthetic capacities in understorey plants is a limited ability to process the light energy they absorb during strong sunflecks. This limited ability can also be exacerbated by a low induction state. Under these conditions, understorey plants will need to dissipate excess energy if they are to avoid photodamage. Field measurements of chlorophyll fluorescence from A. macrorrhiza show a decline in the quantum yield of photosynthesis (measured as Fv/Fm) during saturating sunflecks, indicating that photoprotective mechanisms are probably being engaged. Simultaneous assessment of the xanthophyll pigments shows that interconversion of violaxanthin to zeaxanthin is also occurring. After the sunfleck has passed, conversion of zeaxanthin to violaxanthin is extremely slow in species such as A. macrorrhiza, perhaps allowing a more rapid photoprotective response for subsequent sunflecks. However, quantum yield increases more rapidly than xanthophyll reconversion on return to low light, demonstrating a requirement for both high ΔpH and zeaxanthin for internal photoprotection to occur (Watling et al. 1997b).
Engagement of photoprotective mechanisms by shade-tolerant plants in an understorey environment may seem surprising, but serves to illustrate the extent of spatial heterogeneity in resource availability which is a feature of most habitats.