Where overheating cannot be avoided, plants have developed a range of mechanisms to tolerate high temperatures and to resist the stresses outlined above. Many of these mechanisms have been harnessed by plant breeders to develop more heat resistant crops (for review see: http://www.plantstress.com/Articles/heat_m/heat_m.htm). Each species has a different capacity to respond to heat stress. The degree of heat tolerance tends to follow the species’ native thermal range, with plants irreversibly damaged by temperatures between 30-40 oC termed ‘heat sensitive’ and those only damaged above 40 oC designated as ‘heat resistant’. A key aspect of tolerance to heat stress is the ability to acclimate and the mechanisms described below are typically up-regulated when plants are exposed to moderate heat stress (Figure 14.28).
Fig14.28.png
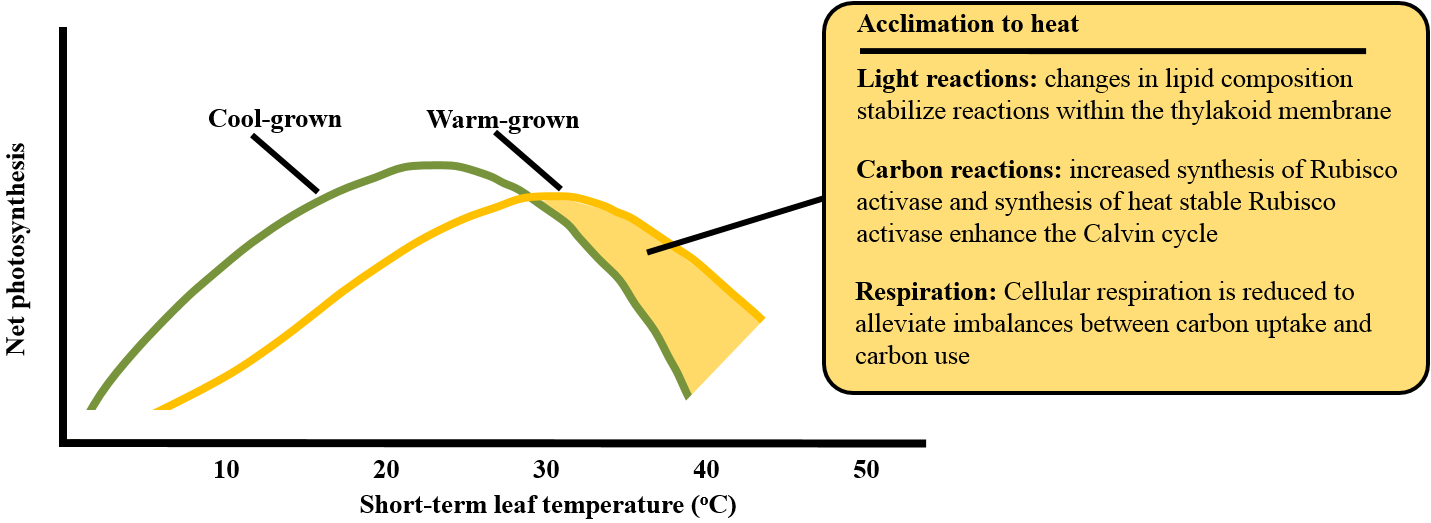
Figure 14.28. In heat tolerant plants, growth at warm temperatures results in acclimation of photosynthesis. Adjustments in membrane composition, protein synthesis, and metabolic regulation alleviate some of the effects of high temperature. Acclimation is mediated, in part, by an increase in expression of heat shock proteins. Based on Sage and Kubien (2007) and Yamori et al. (2014).
(a) Membrane state, structure and composition
In order to tolerate high temperatures, plants must maintain membrane fluidity within a biologically functional range (membrane thermostability). The degree to which membrane fluidity increases with temperature is dependent on membrane composition. Lipids that have unsaturated fatty acid chains, short fatty acid chains or a low sterol content generally form membranes that are more fluid and less stable at high temperatures. The sensitivity of membranes to heat stress can be reduced by increasing the proportion of saturated lipids or by altering the composition of specific lipids. Early work by Jim Lyons and John Raison at the CSIRO Division of Food Research highlighted the fact that tropical species tend to have a higher proportion of saturated lipids than temperate species, but found that the full role of lipid composition in regulating membrane fluidity was complex. Changes in lipid composition during acclimation to high temperature, including increases in the proportion of saturated lipids, have been described in cyanobacteria (Los and Murata 2004) and a number of plants from both warm and cool regions (Raison et al. 1982; Larkindale and Huang 2004). Some of the changes in the physical properties of membranes are regulated by the activity of heat shock proteins (see below), but others are not (Sharkey 2005).
Alteration of lipid composition through gene manipulation has been shown to increase heat tolerance in Arabidopsis, soybean and tobacco (Alfonso et al. 2001; Murakami et al. 2000). Murakami et al. (2000) produced transgenic tobacco plants with a reduced proportion of trienoic fatty acids (unsaturated lipids with three cis double bonds) in the chloroplast membranes. Exposure of the plants to 45oC for 5 minutes reduced photosynthesis by 50 % in the wildtype while the transgenic plants were unaffected (all plants showed complete inhibition of photosynthesis after exposure to 50oC for 5 minutes).
(b) Heat shock proteins
Within minutes of temperature rising above the optimum, the expression of most genes used for general metabolism is inhibited, however, a sub-set of specialised stress response genes are actively up-regulated. The best characterised of these genes are a multi-family group known as heat shock proteins (HSP). HSP occur in all organisms. In plants, they show differential expression in many tissues and many cell compartments. HSP utilise a novel transcription factor to respond directly to heat, and their levels have been shown to rise along with temperature until the lethal threshold temperature is reached. On exposure to high temperature HSP expression typically peaks after 1-2 hours and diminishes after 6-8 hours, after which the cell environment is modified enough for the transcription and translation of other genes to resume (Larkindale et al. 2005; Allakhverdiev et al. 2008).
Many HSP are thought to act as chaperone proteins, protecting other proteins from denaturation by reducing misfolding, unfolding, and aggregation. Chaperone activity also helps maintain the translocation of proteins across cell membranes. The up-regulation of HSP has been shown to improve tolerance to and recover from heat stress in several systems, e.g. addition of purified HSP of isolated tomato chloroplasts significantly improved their heat tolerance by protecting PSII electron transport (Allakhverdiev et al. 2008). This chaperone role can offer protection from stresses aside from heat, and HSP have been shown to be up-regulated by a variety of stresses that escalate protein denaturation. Larkindale et al. (2005), describe five classes of HSP the names indicating the molecular weight:
- HSP60 and HSP70 have been shown to act as chaperone proteins in plants and other organisms, and some also function to stabilise membranes preventing the loss of permeability.
- HSP90 are less well characterised in plants, they are thought to interact with signal transduction proteins that form part to the overall heat stress response.
- HSP100 act as chaperone proteins in conjunction with HSP60 and HSP70 and may perform other roles. Plants lacking HSP 100 can grow normal at optimum temperatures but are unable to acclimate during heat stress.
- Small HSP are particularly important in plants but are less well characterised than HSP60 and HSP70. Small HSP are a diverse group including several gene families that are targeted to different cellular compartments, including the cytosol, chloroplast and mitochondria. The function of many of the Small HSP is still unknown. Some may be involved in chaperone protein activity and some are involved in maintaining membrane stability including the protection of membranes essential for the functioning of PSII.
Certain HSP also act to clean up the cell, removing denatured proteins by increasing the proteolysis activity of ubiquitin. The removal of potentially toxic protein aggregations is thought to be a key part of acclimation to heat stress. Indeed, heat stress also stimulates the up-regulation of ubiquitin itself (Larkindale et al. 2005).
(c) Reactive Oxygen Species (ROS)
The impairment of metabolic function during heat stress results in increased production of ROS, which in turn causes secondary damage to proteins and membranes. Accumulation of ROS during heat stress has been associated with both the light reactions and the Calvin cycle reactions. The reaction centre of PSII is particularly vulnerable, producing superoxide radicals, hydroxyl radicals and hydrogen peroxide under heat stress. Antioxidant enzymes and non-enzyme systems serve to limit the formation of the most damaging ROS, such as singlet oxygen, and to detoxify the cells through ROS-scavenging. Although some antioxidant systems are impaired at high temperatures, others are up-regulated and can be considered part of the heat stress response (Sharkey 2005; Larkindale et al. 2005; Allakhverdiev et al. 2008).
(d) Heat stress response: other mechanisms
Although HSP form a critical part of the heat stress response, they still account for a small minority of the transcripts that are up-regulated during heat acclimation. Several groups are currently working to elucidate the role of other changes in the transcriptome, proteome, metabolome and lipidome in regulating signal transduction and response to heat stress (for review see Mittler et al. 2012). Among the metabolites associated with heat acclimation perhaps the best characterised are the compatible solutes (which also play a crucial role during water stress and salinity stress (see Chapter 17). In the case of heat stress, their primary role is thought to be similar to that of the chaperone proteins, i.e. the protection of protein and membrane function (Larkindale et al. 2005; Allakhverdiev et al. 2008). There is also increasing interest in the role of isoprene in heat tolerance. Isoprene is a small hydrocarbon sometimes released from plants in large quantities. It is particularly associated with certain tree species, such as eucalyptus. Indeed, it is isoprene that is responsible for the distinctive blue haze that characterises Australia’s ‘blue mountains’. Although the full purpose of isoprene has not been established, there is good evidence that it is involved in the development of heat tolerance. In particular, the production of isoprene has been shown to reduce the inhibition of photosynthesis during moderate heat stress, perhaps by associating with the thylakoid membrane to increase hydrophobic interactions and protect membrane function (Sharkey 2005).