Fruit that store starch switch from starch synthesis during development to starch hydrolysis during ripening. Starch–sugar interconversion involves a larger number of enzymes and a greater complexity of control than is required for sugar storage alone. No transgenic plants have yet been reported in which a starch-storing fruit or lipid-storing fruit has been altered to store only sugars, or vice versa, but results with potato suggest it could be possible. In potato, control of both insoluble carbon storage and sugar to starch conversion has been attained using sense and antisense constructs to alter the expression of specific carbohydrate enzymes (Stitt and Sonnewald 1995).
Fig 11.11.png
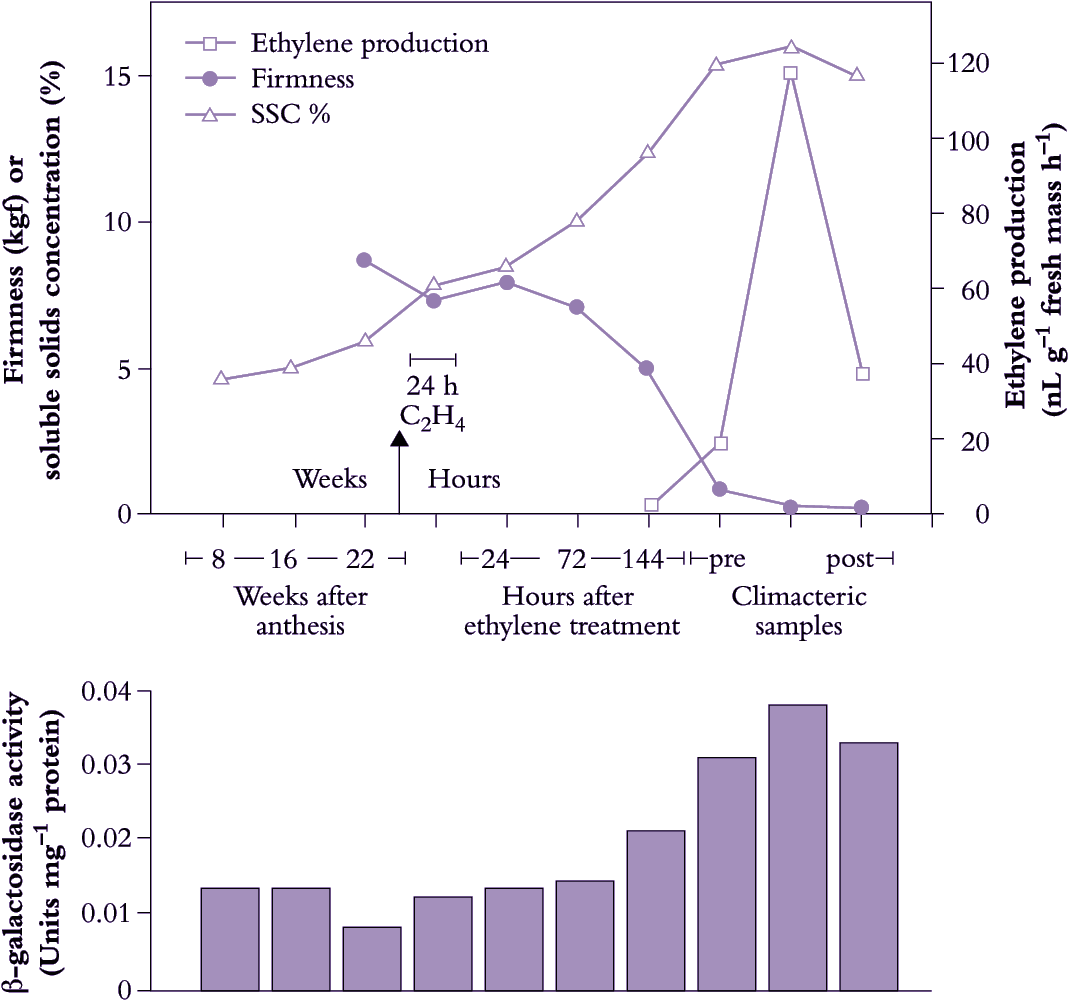
Figure 11.11. Kiwifruit show some dramatic changes in physiological status during development. For 24 weeks after flowering, fruit are hard and sugar content is low. Meanwhile starch content (not shown) rises to about 12% of fresh mass. Ripening in fruit harvested around 24 weeks can be triggered by exposure to an external source of ethylene even though the fruit are not yet producing ethylene by themselves, and are incapable of an autocatalytic response. Within a week of such treatment starch becomes hydrolysed and sugar concentration rises from 7% to 15%. Fruit then soften rapidly and several cell wall-modifying enzymes increase in activity. Fruit do eventually show a peak in ethylene production, but not until ripening is well under way. (Figure based on data from J. Win)
Kiwifruit provide an example of biochemical changes in a starch-storing fruit (Figure 11.11). They are normally harvested with starch contents ranging from 4 to 10% (dry matter concentrations between 14 and 20%; SSC between 6.2 and 12%). At harvest, the main sugars are sucrose, glucose and fructose. As fruit ripen after harvest, sucrose content increases only slightly, while fructose and glucose increase in parallel to become the predominant sugars in ripe fruit. Labelling with radioactive precursors indicates that all three sugars are actively synthesised during ripening, and there are increases in the activities of a number of sucrose-metabolising enzymes, particularly SPS and invertase. The starch-degrading enzyme α-amylase increases two-fold.
The organic acid content of kiwifruit also changes after harvest. At room temperature malate decreases and citrate increases while quinate remains unchanged. However, during cool storage (0–4°C), malate increases. Such changes are enough to alter the flavour balance in the ripe fruit. Where fruit store carbon in an insoluble form, there are several potential control points for sugar metabolism (Figure 11.9) including:
- Hydrolysis of starch to glucose;
- Transfer of sugar precursors from starch-containing plastids (amyloplasts);
- Synthesis or degradation of sucrose;
- Synthesis of hexoses;
- Transfer of sugar to vacuoles or export from cells;
- Carbon flow between sucrose and malate or citrate;
- Production of CO2 from sugar or acid precursors;
- Transfer of malate or citrate across the vacuolar or mitochondrial membranes.
Interference in any of these processes should affect ripening and/or flavour development after harvest. Such interference may be physical (as in storage temperature), chemical (as in atmosphere composition) or genetic (by modifying activities of specific proteins which control flow between particular metabolites).