Fig_18.12.png
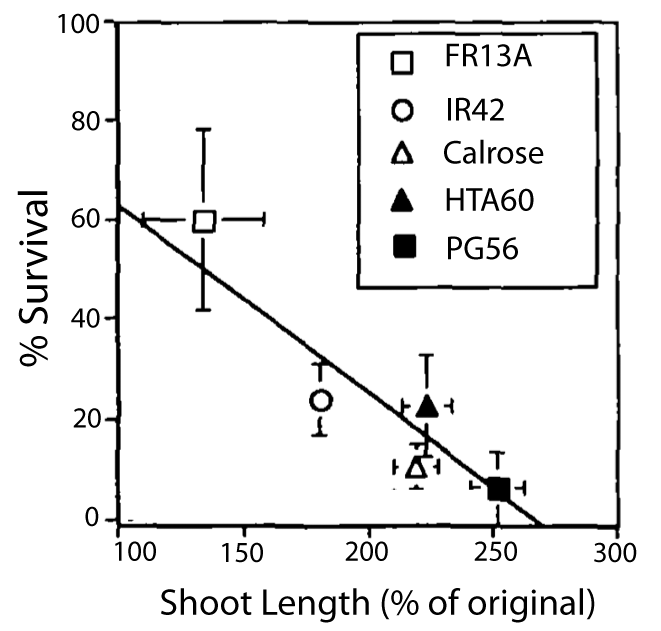
Figure 18.12. Shoot elongation of rice cultivars during submergence relative to survival. Plants were submerged 14 d at a mean daily irradiance of 22.3 ML m-2. Open symbols, lowland rice; closed symbols, deepwater rice cultivars. Setter and Laureles (1996).
Leaves, petioles and stems of completely submerged herbaceous plants are generally not anoxic, although can become hypoxic. The internal O2 concentration in these submerged shoot organs is determined by the rate of photosynthesis under water, the rate of respiration of the tissue and the rate of gas exchange with the external water medium. This results, for example, in endogenous petiole O2 concentrations in fully submerged Arabidopsis thaliana of 17 kPa during the light period and 6 kPa during darkness (Lee et al. 2011; Vashisht et al. 2011). Due to the strong variation over time, O2 is seen as an unreliable indicator of submergence of shoot organs. Therefore, shoots make use of the entrapment of another gaseous component, the plant hormone ethylene, to sense the change of the outside environment from air to water. Similar to other gases, ethylene only very slowly diffuses in water. Since ethylene is produced by every plant cell, slow diffusion in water leads to a substantial increase of ethylene levels inside shoot tissues within less than one hour of submergence. This enhanced endogenous ethylene concentration is guaranteed in submerged shoots as long as some O2 is present to maintain the O2-dependent ethylene biosynthesis.
Flood tolerant plants exposed to complete submergence exploit two contrasting suites of traits, escape or quiescence, to survive this stress. In brief, plants with the escape strategy: (i) increase the growth rate of shoot organs, such as petioles and stems, so as to emerge above floodwaters, and (ii) initiate the development of aerenchyma to facilitate internal gas diffusion. Quiescent plants, on the other hand, “wait out the submergence event” and are characterized by: (i) conservation of energy and carbohydrates via, for example, a reduction of the underwater growth rate, and (ii) an increase of molecular components that prepare shoot and root organs for future conditions with low O2 and production of protective molecules that counteract harmful cellular changes associated with flooding, such as production of ROS.
A classical study with a range of rice cultivars revealed that shoot elongation under water without reaching the surface goes at the expense of survival and thereby demonstrating elongation is associated with costs (Figure 18.12). From an evolutionary point of view the escape strategy will only persist if these costs are outweighed by benefits such as improved aeration, energy generation and carbon production, and ultimately improved survival, growth and reproduction. Therefore, the escape strategy is restricted to plants in environments with shallow floods or with deeper floods that persist over longer durations as the case with deepwater and floating rice. Transient very deep or ephemeral floods, however, favour plants with the quiescence strategy.
Fig_18.13.png

Figure 18.13. Petiole elongation response in Rumex palustris upon submergence. Plant on the left-hand-side was in air, plant on the right-hand-side was submerged for the final 10 days. (Photograph courtesy of Liesje Mommer).
Fast extension of shoot organs in response to submergence is described for species from a wide range of families. As an example, Figure 18.13 illustrates submergence-induced petiole elongation in the semi-aquatic plant Rumex palustris.
Depending on the tissue type and the developmental stage of the shoot organ, fast underwater growth involves cell elongation only (e.g. petiole of Rumex palustris) or a combination of increased cell division and elongation (e.g. stem of deepwater rice). Fast cell elongation is regulated by specific transcription factors and by an interacting set of plant hormones. In deep water rice, characterized by an enormous stem elongation capacity upon submergence, ethylene regulates two important genes, SNORKEL1 and SNORKEL2, which encode nuclear-localized DNA binding proteins that belong to the Ethylene Response Factor (ERF) family of transcription factors. Non-deepwater rice varieties lack these ERF genes and their importance for elongation was demonstrated by introgression of these loci from deepwater rice into non-elongating varieties which then showed substantial elongation when submerged. Next to ethylene, three other downstream-operating plant hormones are involved in submergence-induced shoot elongation. Upon submergence, levels of abscisic acid are quickly reduced, whereas auxin and gibberellic acid increase. Ultimately these signal transduction components affect the rate limiting step for cell elongation: the cell wall. In order to allow turgor-driven cell expansion, cell walls must loosen by means of specific cell wall loosening proteins such as Expansins. The expression of Expansin genes is strongly upregulated and the abundance of Expansin proteins increases, soon after submergence of species with shoots that elongate.
An important trait for plants that survive flooding by means of the quiescence strategy is reduction of underwater growth to conserve carbohydrates and retention of chlorophyll to enable continued, albeit reduced, photosynthesis. Increased submergence tolerance in rice caused by reduced plant growth rates under water is regulated by the ethylene-induced expression of the SUB1A-1 gene. Interestingly, this gene belongs to the same ERF transcription factor family as the two SNORKEL genes. SUB1A-1 limits elongation growth by two mechanisms: (i) minimizing the decline in the gibberellin signaling repressor SLENDER RICE-1 and the related SLENDER RICE LIKE-1, and (ii) enhancing GA catabolism by differentially regulating genes associated with brassinosteroid synthesis in submerged shoots (Schmitz et al. 2013). On top of that, SUB1A-1 also inhibits synthesis of ethylene, expression of Expansins and reduces starch and sucrose reserve depletion. Recently, SUB1A-1 was crossed into high yielding rice varieties leading to more flood tolerant varieties that have recently been released to farmers in Asia. These varieties have yield advantages of 1 to over 3 tons/hectare over the varieties lacking SUB1A-1 following submergence for various durations (Mackill et al. 2012). Figure 18.14 demonstrates increased survival and yield after 15 d of complete submergence during the vegetative stage followed by recovery after de-submergence of a rice variety containing SUB1A-1 compared with the original variety that lacks it.
Fig_18.14.png

Figure 18.14. IR64 (left) and IR64-Sub1 (right) after 15 days of submergence during vegetative stage in the field. Two-week-old seedlings were transplanted into a field, grown for another two weeks then completely submerged for 15 days. The field was then drained and plants were allowed to recover under non-stress conditions. The photograph was taken about 90 days after de-submergence. (Photograph courtesy of AM Ismail).
In summary, most plants cannot withstand complete submergence lasting over a few days; however, semi-aquatic plants such as certain rice genotypes can survive complete submergence even for over two weeks. Tolerance of rice to transient submergence is mainly achieved by restricting growth and respiration, thus conserving carbohydrate reserves to enhance recovery when the floodwater recedes.