Fig6.16_edit.png
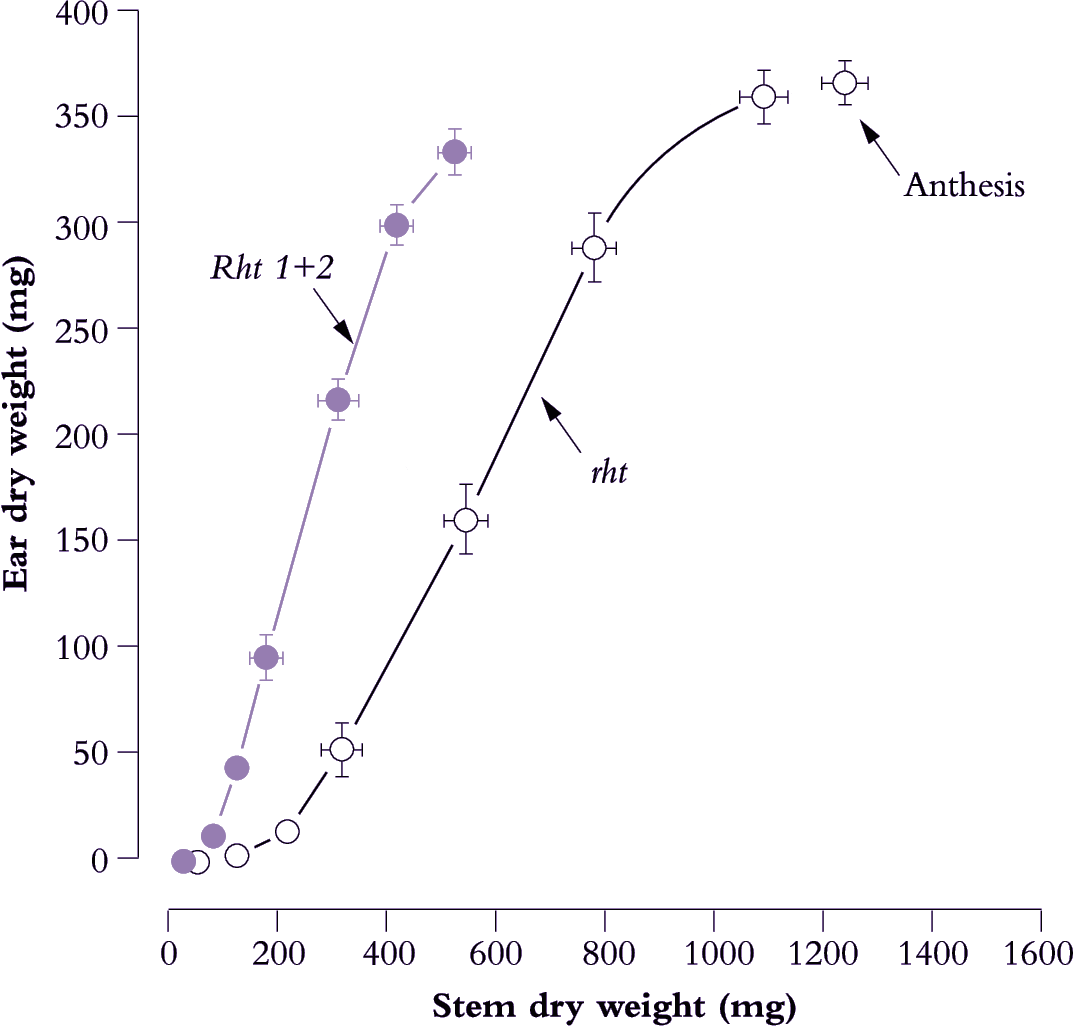
Figure 6.16. Early growth of reproductive tissues relative to stem mass in dwarf genotypes foreshadows faster ear development and higher HI. The tall and productive Mexican spring wheat (Yaqui 50, designated rht) eventually produces heavier ears, but returns a lower HI at maturity. Introduction of two major dwarfing genes (Rht 1 + Rht 2) resulted in shorter stems. Consequently developing ears were subject to less competition for photoassimilate during early differentiation and for grain filling subsequent to anthesis. Bars represent standard errors. Based on Bush and Evans (1988) Field Crops Res 18, 243-270
A major impetus to improve HI in cereals came from the introduction of dwarfing genes. In primitive wheats, and tall plants generally, reproductive structures have to compete with rapidly extending stems for photoassimilate, but dwarf cultivars alleviate such competition and enable a shift in carbon partitioning to ears. Early growth of ears and stems in two lines of a Mexican spring wheat (Figure 6.16) illustrate this principle. A steeper slope in the dwarf line (designated Rht 1+2) compared with the tall line (rht) implies greater allocation of photoassimilate to ear growth relative to stem growth. The two dominant dwarfing genes (Rht 1 plus Rht 2) which are insensitive to gibberellic acid result in short stems and enhanced yield. Such genotypes formed the basis of the Green Revolution.
Tall wheat commonly “lodges” (falls over) in nitrogen-rich conditions, and dwarf wheats were originally developed to overcome this problem. Crop physiologists and breeders subsequently recognised the yield advantage from improved partitioning of photoassimilate. Continuing selection for yield within the semidwarf background, which became common after the 1970s in both spring and winter wheats, has seen further yield progress with little change in plant height. Yield has improved further in spring wheats (Sayre et al 1997; Sadras and Lawson 2011) and in winter wheats (Shearman et al 2005) but yield progress in more recent varieties is associated with notable increases in total biomass. This has occurred in both spring wheats (Sadras and Lawson 2011) and winter wheats (Shearman et al 2005; Zheng et al 2011). Also whereas past yield progress in wheat has always been associated with more grains m-2 and unchanged or smaller grains, yield gain in recent varieties in some cases has been linked to heavier grains (e.g. Zheng et al 2011). The switch from higher harvest index to greater biomass may reflect limits to harvest index, now around 0.5 for the best varieties about 80-100 cm in stature, while the appearance of newer varieties with heavier grains may reflect pressure from grain processors. Either way the changes point to the power of empirical selection.
Some room still exists for further improvement in shoot HI compared with 1980s values (Figure 6.15) but there is a corollary. If shoot biomass remains unchanged, further improvement in HI implies further reduction in leaf and stem mass. Considering leaves, specific leaf area (area:mass ratio or SLA) will have a finite limit for structural reasons so that the area of CO2-assimilating tissue servicing those enlarged sinks must also reduce as mass is reduced. Net assimilation per unit area (NAR) will therefore need to increase even further if potentially higher yields are to be realised. This could be achieved by either increases in photosynthetic potential or in more efficiency use of the energy produced.
The following section deals with respiratory efficiency and plant growth.