3.0-Ch-Fig-3.11d.jpg
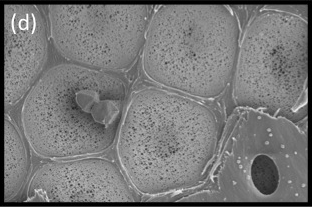
Figure 3.11d SEM showing surface view of the pit membrane with secondary wall removed from most pits by sectioning. Tiny pores allow the movement of water between vessels but limit the movement of gas and pathogens. Bordered pits act as the safety valves of the plant hydraulic system. (Photograph courtesy B Choat and S Jansen)
Xylem vessels (and tracheids) are not just pipes to carry water, they are pipes with holes in them (pits) through which water can leak out, fulfilling a principal leaf function of water distribution through the transpiration stream to places where it will evaporate. The branching network of vessels is beautifully adapted to achieve this.
Think about flow in a leaky vessel. As explained earlier, the rate of volume flow varies as the fourth power of the radius (Poiseuille’s Law). The frequency of leaks through vessel walls varies with surface area of the walls, that is, as the first power of the radius (2πr). So if the vessel is wide, forward flow is much larger than leakage. The wide vessels in large veins supply water all over the leaf without losing much on the way. As the width of a vessel becomes smaller, the forward flow (a function of r4) is reduced much more strongly than the leaks (a function of r). The proportion of water lost to leakage therefore increases as vessels become smaller. Indeed, for a fixed pressure gradient there is a critical radius at which all water entering the vessels supplies leaks, and there is no forward flow at all. The finest veins of dicotyledenous leaves have vessels of a radius that is close to this critical value. As sap disperses into the fine ramifications of the network, it moves more and more slowly forward, and leaks increasingly outwards through the sheath to the mesophyll. This is the rationale of the distributing networks of the small branched veins of both dicotyledons and grasses.
3.0-Ch-Fig-3.29.jpeg
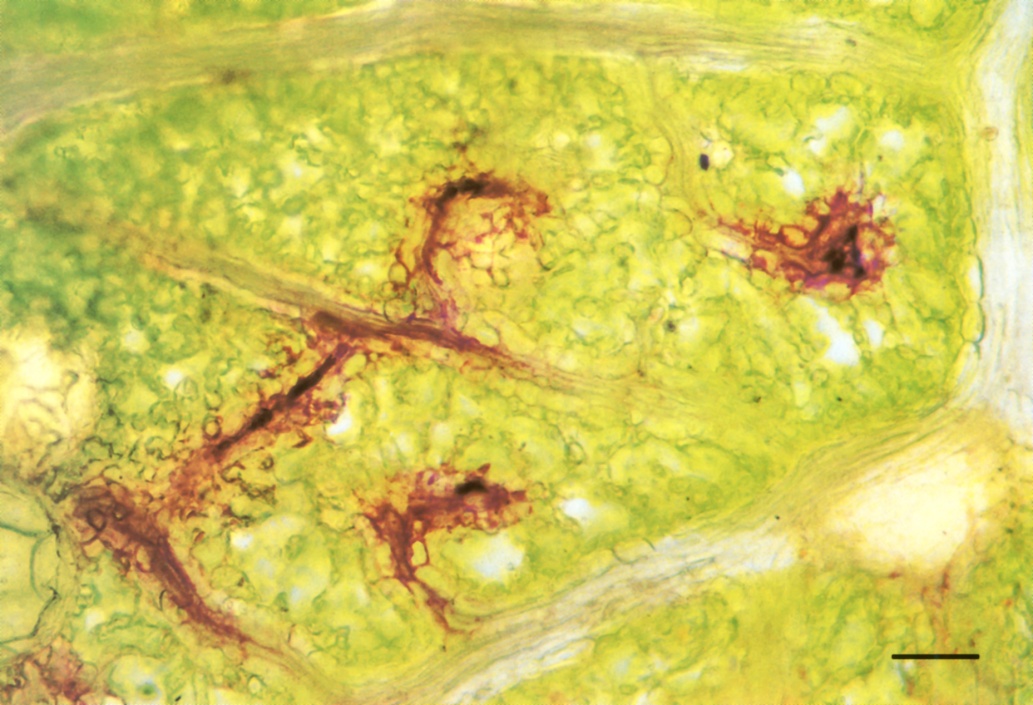
Figure 3.29 Fresh paradermal hand-section of a leaf of Eucalyptus crenulata which had been transpiring for 80 min in a solution of sulphorhodamine G. The dye solution is present at low concentration in the vessels of the larger veins, but is not visible at that concentration. In the smallest veins the dye has become so concentrated by loss of water to the symplasm that dye crystals have formed inside vessels. Sectioned in oil, bright-field optics. Scale bar, 100 µm. (Photograph courtesy M. McCully and M. Canny)
Extraction of water from fine veins can be readily demonstrated. Stand a cut leaf in an aqueous solution of dye, such as 0.1% sulphorhodamine G, and allow it to transpire for an hour. The solution moves rapidly through large veins all over the leaf in a few minutes. Then it moves increasingly slowly into the network of distributing small veins. By the end of an hour it has reached the ends of the finest veins and, as water is extracted from them, the dye becomes more and more concentrated (Figure 3.29). Movement of dyes from the finest veins to leaf surfaces 100 µm away takes about 30 min, suggesting that diffusion rather than mass flow is responsible for solute distribution to cells.