The simplest vein architecture is found in conifer needles where a single unbranched strand of xylem and phloem is surrounded by mesophyll (Figure 3.22).
3.0-Ch-Fig-3.22.jpeg
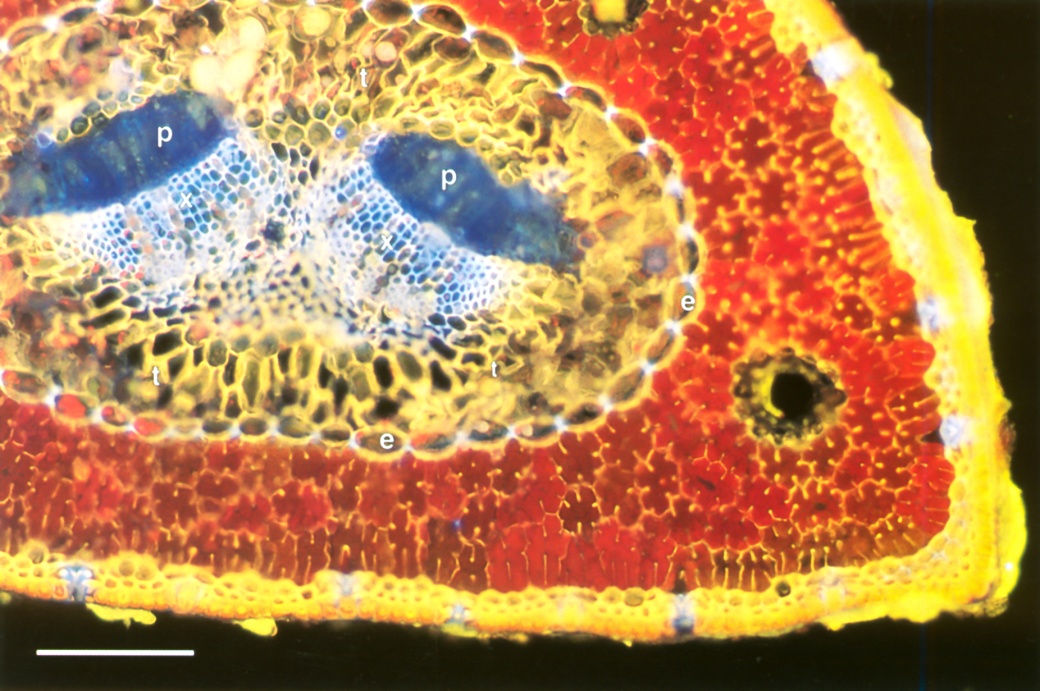
Figure 3.22 Transverse section of a pine needle. A single central vein has two strands of phloem (p) and xylem (x), embedded in transfusion tissue (t). These vascular tissues are separated from the chlorophyll-containing mesophyll (red) by an endodermis (e) with Casparian strips and suberised lamellae. Stomata in the epidermis appear bluish. Rhodamine B stain, fluorescence optics. Scale bar, 0.1 mm. (Photograph courtesy M. McCully)
3.0-Ch-Fig-3.23.jpeg

Figure 3.23 Wheat leaf showing one large supply vein and three small distribution veins, connected by transverse veins. Small veins could carry two thirds of the evaporated water. Dark-field optics, blue filter. Scale bar, 0.1 mm. (Photograph courtesy M. McCully)
Vascular strands are enclosed by an endodermis that separates them from the mesophyll, and are embedded in a mixture of parenchyma cells and tracheids called transfusion tissue. Water from the xylem permeates radially outward through transfusion tissue, endodermis and mesophyll to evaporate below lines of stomata in the epidermis.
Leaves of angiosperms have much more complicated venation than conifer needles. If you look at a grass leaf with your hand lens, parallel veins run the length of the leaf, but they are not all the same size. A few large veins have several small veins lying between them. On closer inspection with a light microscope, all these parallel veins are connected at intervals by very small transverse veins (Figure 3.23).
There are in fact two vein systems with different functions: large veins supply water rapidly to the whole length of a leaf blade while small veins and their transverse connections distribute water locally, drawing it from the large veins. Water in large veins flows only towards the tip, but in small veins it can flow either forwards or backwards along the leaf blade or transversely between adjacent parallel veins. The distinction of flow patterns in large and small veins arises as a result of different vessel sizes. Large veins have wide vessels (about 30 µm diameter), while small veins have narrow vessels (about 10 µm diameter). As the volume of water flowing along pipes is proportional to the fourth power of the radius (Equation 6; Poiseuille’s Law), volume flow in the larger vessels will be 3 to the fourth power (= 81) times greater than the flow in the smaller vessels. Put another way, pressure gradients along the leaf in large vessels will be very slight, but steep pressure gradients can develop locally in narrow vessels that will direct local flows into the mesophyll. Large veins supply water rapidly over the whole lamina while small veins distribute it locally and slowly. The slower flux along minor veins is compensated by their far greater number, which results in them having a greater length per unit leaf area than major veins.
With a hand lens, you do not see the vessels, instead you see the sheaths that surround xylem and phloem, much as an endodermis surrounds vascular strands of a conifer needle. Grass leaf veins have two sheaths of cells surrounding the parallel veins and containing the xylem and phloem tissues. In the leaf anatomical development typical of C3 species (e.g. wheat), both sheaths are parenchymatous and lack chloroplasts (Figure 3.24).
3.0-Ch-Fig-3.24.jpeg

Figure 3.24 Wheat leaf showing a single large (supply) vein comprising three large vessels (V). The vein is surrounded by two sheaths of living cells, the inner mestome sheath (arrowheads), and outer parenchyma sheath (stars). The mestome sheath of these veins is impermeable to water. Water enters the symplasm at the inner boundary of the parenchyma sheath (Canny 1990). Phloem (P). Transverse hand-section, Toluidine blue stain, bright-field optics. Scale bar, 0.1 mm. (Photograph courtesy M. McCully and M. Canny)
3.0-Ch-Fig-3.25.jpeg
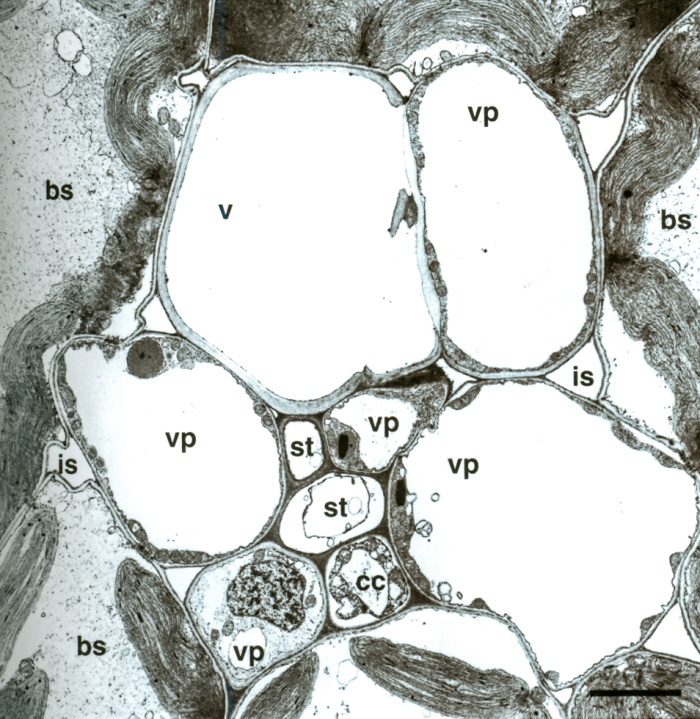
Figure 3.25 Minor vein of maize, consisting of one xylem vessel (v), five vascular parenchyma cells (vp), bundle sheath (bs), companion cell (cc), and sieve tube (st) with intercellular space (is). Scale bar, 4.2 µm. Transmission electron micrograph, R.F. Evert. (Reproduced from Planta 138: 279-294, 1978)
The large veins of a C3 species is surrounded by two sheaths of living cells, the inner mestome sheath and outer parenchyma sheath (Figure 3.24). The mestome sheath of these large veins is impermeable to water. There is no apoplastic path for water through the mestome sheath of large veins, except through a connecting transverse vein. In small veins, by contrast, two or three mestome sheath cells next to the xylem permit flow of water and solutes through the cell wall apoplasm.
In C4 species (e.g. maize), only the inner (mestome) sheath is without chloroplasts. The outer ring of sheath cells contains large chloroplasts and is known as the bundle sheath (Figure 3.25). This is the cell layer in which CO2 fixation in the Calvin cycle takes place in C4 plants. This is an essential part of the carbon concentrating mechanism, and the special anatomy of C4 photosynthesis, as detailed in Chapter 2, Section 2.2.2
A dicotyledonous leaf contains the same two vein systems as a grass leaf, but these are differently arranged. Large supply veins are prominent, comprising a midrib and two orders of branches off it, often standing out from the surface of the lamina. These contain wide vessels and carry water rapidly to the leaf margins. Between them lie distribution veins, another two branch orders of small veins dividing the mesophyll up into islets about 1–2 mm across (Figure 3.26), and within these islets a fifth and final order of branches of the finest veins. The fourth- and fifth-order branches have only narrow vessels (Figure 3.26).
In dicotyledenous leaves, as in grass leaves, these vascular tissues are enclosed by bundle sheath cells through which water and solutes must pass when leaving the xylem.
3.0-Ch-Fig-3.26.jpeg
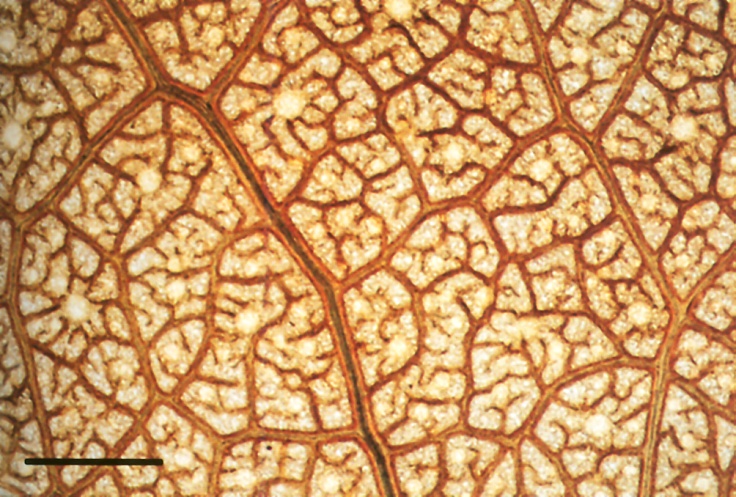
Figure 3.26 Whole mount of a cleared leaf of Eucalyptus crenulata showing the complicated arrangement of supply and distribution veins characteristic of a dicotyledonous leaf. Islands marked out by large veins with large vessels, in which water is moved rapidly all over the lamina, surround islets of small veins with small vessels in which water is slowly distributed locally to the mesophyll. Partial phase-contrast optics. Scale bar, 1 mm. (Photograph courtesy M. McCully and M. Canny)
Any distribution network such as the branching vessels of decreasing size in leaves is found to obey Murray’s Law. This states that the cube of the radius of a parent vessel is equal to the sum of cubes of the radii of the daughter vessels (e.g. a 50 µm vessel would branch into five 30 µm vessels). Such a pattern of branching produces optimal flow in several senses: minimum energy cost of driving that flow, minimum energy cost of maintaining the pipeline, constant shear stress at the walls of pipes, and rapid flow in supply pipes with slow flow in distributing pipes to permit exchange through the pipe walls (LaBarbera 1990).
3.0-Ch-Fig-3.27.jpeg
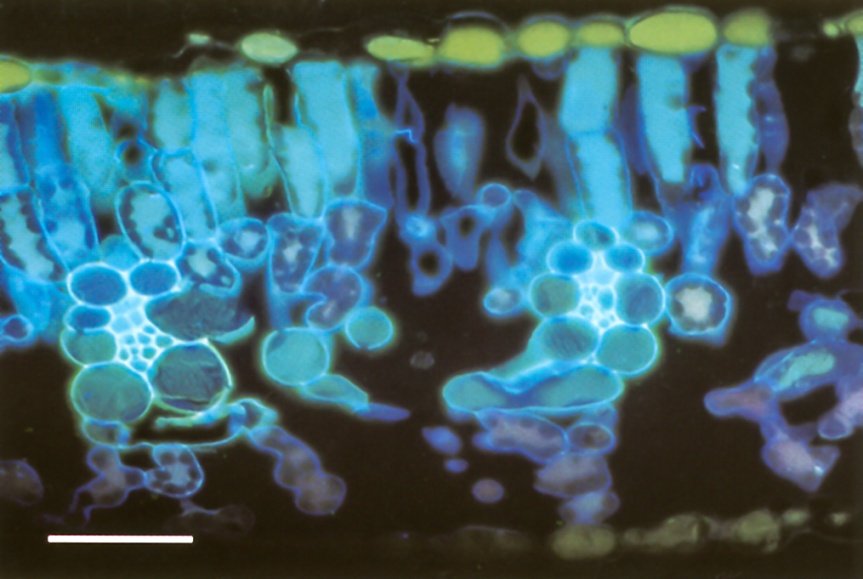
Figure 3.27 Soybean leaf showing two of the smallest veins surrounded by bundle sheath cells. Veins of this size are the end of the branching network shown in Figure 3.26, and supply most water that is evaporated. This leaf was transpiring in a solution of fluorescent dye for 40 min. The small vessels in each vein contain dye solution which has become concentrated by water loss to the symplasm and out through the bundle sheath. Dye has started to diffuse away from small vessels in the cell wall apoplasm of bundle sheath cells. Anhydrous freeze-substitution and sectioning, fluorescence optics. Scale bar, 50 µm. (Photograph courtesy M. McCully and M. Canny)
The ring of cells forming the sheath around the xylem and phloem tissue (Figure 3.27) acts both as a mechanical barrier that may confine pressure within the vein, and a permeability barrier that can control rates and places of entry and exit of materials. Exceptions are to be found at the ultimate ends of some dicotyledonous fine veins where tracheids or sieve elements may be unaccompanied by other cells, in the transverse veins of grasses which have no sheath, and in some special veins at leaf margins where the sheath is absent on the xylem side. There is evidence of a suberised layer in walls of these sheaths cells in some species, but not in others. It is uncertain whether xylem sap must traverse the cell membranes, as in the suberised endodermis of roots, or if it can travel along the cell walls.