3.0-Ch-Fig-3.33.jpg

Figure 3.33 Leaf of horse chestnut, Aesculus hippocastanum, showing as with all strongly-toothed leaves, conspicuous veins running to each tooth. (Photograph courtesy M. Waters)
(a) Phloem export
Potassium is known to be extracted from the xylem and re-exported via the phloem (Section 5.1). In leaves, the concentrated sap flowing through the narrow xylem vessels of fine veins is separated from sieve tubes of the phloem by only two or three parenchyma cells, frequently enhanced by transfer cells. Exchange from xylem to phloem is probably made by this direct path. Solutes travel in the phloem back to stems, either distally to younger developing leaves and the stem apex or proximally towards roots.
(b) Scavenging cells
Recycling of solutes out of leaves can also be fed by a variety of special structures adapted for processing rather larger volumes of sap, and so suited for collecting solutes present in the stream at quite low concentrations. The transfusion tissue of conifer needles is one of these structures. As sap leaves the xylem of a vascular strand it moves through a bed of tracheids mixed intimately with transfusion parenchyma cells (Figure 3.22 shown earlier). The endodermis acts as the ultrafiltration barrier, allowing water to pass through while leaving dilute solutes to accumulate in transfusion tracheids. Transfusion parenchyma cells have very active H+-ATPases in their cell membranes which accumulate selected solutes (certainly some amino acids) back into the symplasm for return to the phloem and re-export. Such actively accumulating cells are called scavenging cells.
A tissue that acts in the same way is a special layer of cells in the central plane of many legume leaves (extended bundle sheath system or paraveinal mesophyll). It consists of scavenging cells with active H+-ATPases and accumulates amino acids, stores them and forwards them to developing seeds via the phloem.
Jagged ‘teeth’ on the margins of many leaves also contain scavenging cells (Figure 3.33). Veins carry large volumes of the xylem sap to these points, where evaporation is especially rapid. Within a ‘tooth’, xylem strands end in a spray of small vessels among a bed of scavenging cells. Scavenging cells can thereby collect amino acids and load them into the phloem.
(c) Solute excretion
Not all the solutes of the transpiration stream are welcome back in the plant body. Some, such as calcium, are immobilised in insoluble compounds (calcium oxalate crystals) and shed when leaves fall. Others are excreted through the surface of living leaves. A striking excretion system is found along the margin of maize leaves. Here the outermost vein has a single very wide vessel. Rapid evaporation from the exposed leaf edge cooperates with this low-resistance vessel to draw to the leaf margin all residual solutes that have not been taken out of the stream by other veins. Thus foreign material (like dyes) accumulates in this outermost vessel. The vein sheath is missing from the outer part of this vein so that vessels abut directly the airspace at leaf margins (Canny 1990). Solutes are excreted from this marginal vein, dissolved out by rain and dew, and, more actively, at night time by guttation fluid when there is positive pressure in the xylem. A similar accumulation of dye from the transpiration stream is shown along the margin of a eucalyptus leaf in Figure 3.34.
3.0-Ch-Fig-3.34.jpeg
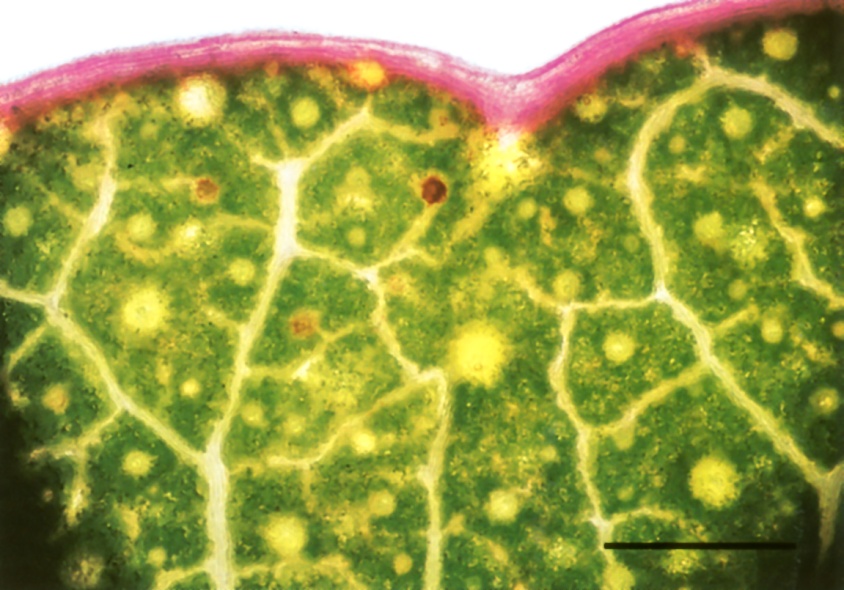
Figure 3.34 Whole mount of the living margin of the eucalypt leaf sectioned in Figure 3.11 prepared after dye had been fed to the cut petiole for 90 min. Dye has spread to the leaf margin in large veins where it accumulates at high concentrations. By analogy with maize leaves (Canny 1990) this is likely to be a system for excreting unwanted solutes. Bright-field optics. Scale bar, 1 mm. (Photograph courtesy M. McCully and M. Canny)