More than 40 eudicot and monocot species distributed over 21 lineages have been reported to possess intermediate C3 and C4 photosynthetic characteristics and CO2 compensation points. These intermediate species are likely remnants of the complex processes that led to the evolution of C4 plants from C3 ancestors, although reversions from the C4 condition have also been suggested. Moreover, a number of identified C3-C4 species occur in taxa that are not closely related to any C4 lineage, raising the possibility that the C3-C4 photosynthetic pathway may be a distinct adaptation. This, in addition to the small number of intermediate species found so far cast doubts over their physiological and ecological fitness, and whether they represent living fossils of evolutionary paths or evolutionary dead-ends (Rawsthorne 1992; Sage et al. 2011).
Leaves of all C3-C4 intermediates have partial or full kranz anatomy, with prominent bundle sheath cells containing chloroplasts and other organelles, and intermediate interveinal distances. Bundle sheath chloroplasts contain Rubisco and functional PCR cycle in both mesophyll and bundle sheath cells. Intermediate leaves also have CO2 compensation points that are lower than what is observed for C3 leaves and can be indistinguishable from C4 leaves, due to reduced photorespiration. Biochemically, C3-C4 intermediates differ in the level of activity of the C4 cycle and the extent to which CO2 is concentrated in bundle sheath cells.
Picture4-1.png
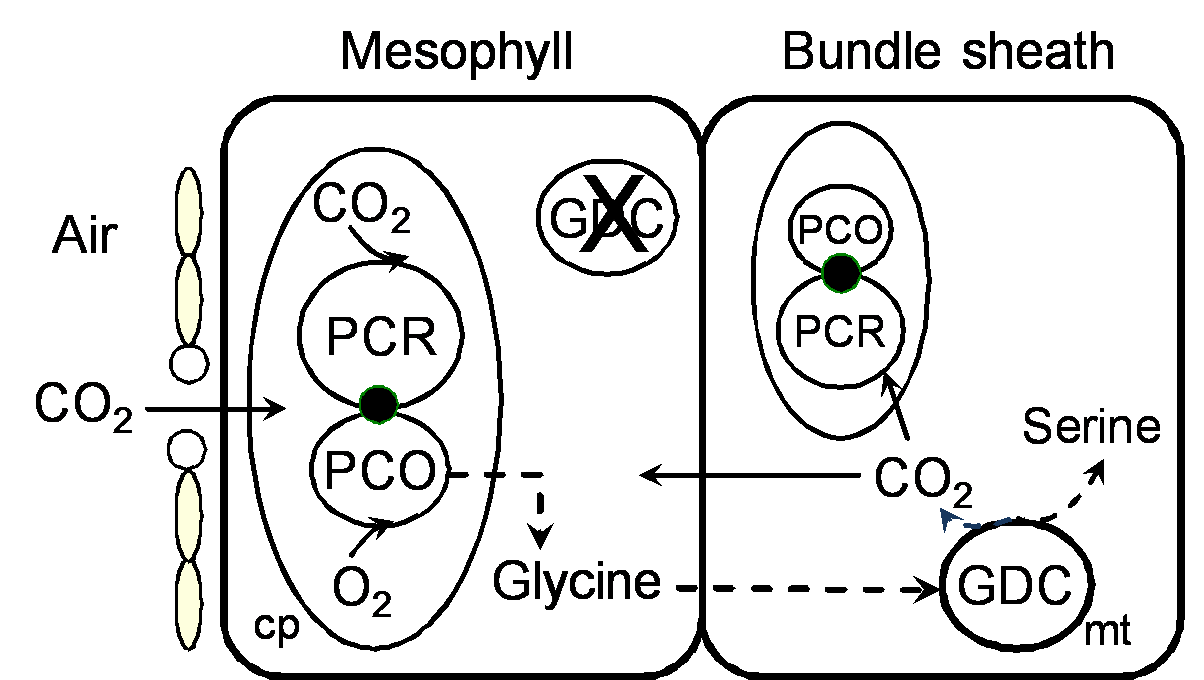
Figure 2.10. Schematic representation of the ‘photorepiratory pump’ operating in C3-C4 photosynthesis. The intermediate photosynthetic pathway reduces photorespiration by refixing photorespired CO2 released locally in the bundle sheath cell. Mesophyll mitochondria lack glycine decarboxylase activity. Mesophyll and bundle sheath cells contain chloroplasts with functional Calvin cycle. Abbreviations: cp: chloroplast; GDC: glycine decarboxylase; PCO: photosynthetic oxidative cycle; PCR: photosynthetic reductive cycle; mt: mitochondrion.
C3-C4 intermediate plants reduce photorespiration (and hence, CO2 compensation point) using a ‘photorespiratory pump’ based on modified localisation of the mitochondrial photorespiratory enzyme, glycine decarboxylase (Figure 2.10). In these plants, glycine decarboxylase activity is restricted to bundle sheath cells and excluded from mesophyll cells. Consequently, photorespired CO2 is released in the bundle sheath where it is largely refixed by Rubisco and the bundle sheath PCR before it diffuses back to the mesophyll. Such a system may weakly elevate CO2 in bundle sheath cells. Intermediate species that rely on the ‘photorespiratory pump’ are termed C3-like or Type I intermediates (e.g., Panicum milioides, Flaveria pubescens) and have intermediate CO2 compensation points and negligible C4 cycle activity.
In Type II and C4-like intermediates (e.g., Flaveria brownii), up to 70% of atmospheric CO2 may be first fixed into C4 acids. These plants have C4-like CO2 compensation points but are not classified as C4 plants because they lack the strict localisation of photosynthetic enzymes (e.g., Rubisco is present in mesophyll cells) and their bundle sheath cell walls have high CO2 permeability, resulting in only a partial CO2 concentrating mechanism (Brown 1980; Ku et al. 1991; Rawthorne 1992; Vogan and Sage 2011).
The physiological advantages of the intermediate photosynthetic pathway in all its naturally occurring forms remain unclear. It may be hypothesised that lowered photorespiration may lead to reduced CO2 limitation of photosynthesis, and thus allow C3-C4 plants to operate with lower stomatal conductance, thus conferring higher water use efficiency relative to C3 counterparts. Moreover, increased nitrogen cost associated with ‘building’ another set of photosynthetic cells (bundle sheath) may reduce nitrogen use efficiency if the gains in CO2 uptake are not substantial.
Work conducted with C3-C4 species yielded inconclusive evidence on the likely advantages of C3-C4 photosynthesis relative to the ancestral C3 mode. Generally, these studies demonstrated that, short of substantial C4 cycle activity and advanced cell-specific localisation of C3 and C4 cycle enzymes between the mesophyll and bundle sheath cells, C3-C4 photosynthesis does not improve photosynthetic efficiency (Bolton and Brown 1980; Pinto et al. 2011, Vogan and Sage 2011). Therefore, partial recycling of photorespired CO2 or a partial CO2 concentrating mechanism reduce photorespiratory loss normally associated with C3 photosynthesis, without leading to significant gains in plant fitness or productivity.