Mary Byrne, School of Biological Sciences, University of Sydney
Leaves are the main photosynthetic organs of vascular plants, and their optimisation for conversion of light to chemical energy production has resulted in a striking array of shapes. There is enormous variation in the shape of leaves of different plant species, of leaves within species, and for some species there is variation between juvenile and mature leaves on an individual plant. In the most general type of dicot leaf, there is a basal stalk or petiole and a distal flat blade or lamina. The leaf forms this structure through development in three axes; the proximal-distal (tip to base), the adaxial-abaxial (top to bottom) and the medial-lateral (middle to margin) (Figure 7.11). The adaxial-abaxial axis is evident at early stages of development of the leaf primordium. The adaxial side of the leaf is the side nearest to the shoot meristem and the abaxial side of the leaf is the side further from the meristem (Figure 7.11). These will eventually become the upper and lower sides of the leaf.
7.1-Ch-Fig-7.11.jpg
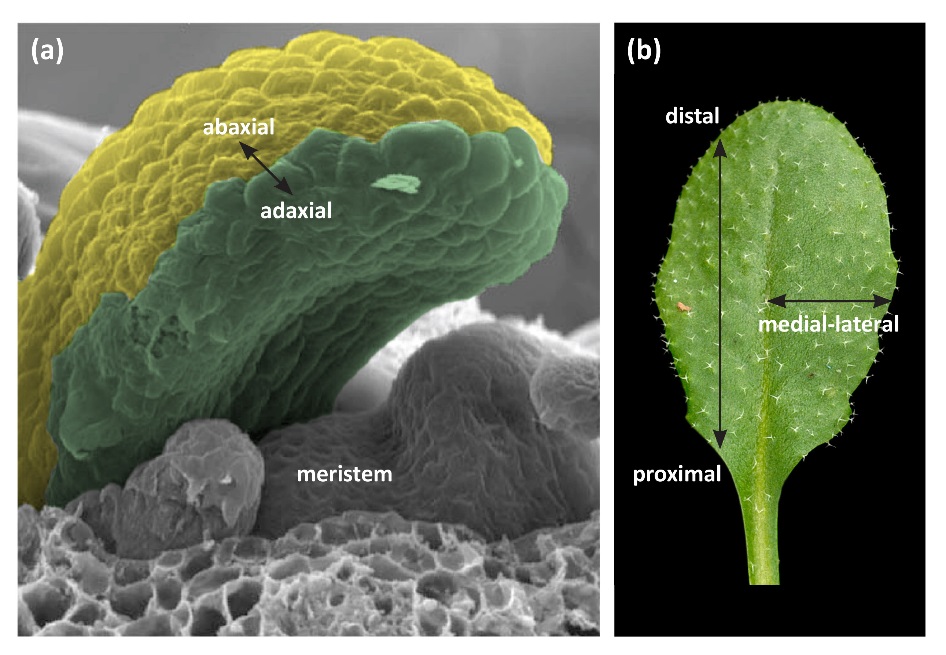
Figure 7.11 Diagram of a plant apex and mature leaf of a dicot such as Arabidopsis. (a) The shoot meristem comprises a population of undifferentiated dividing cells. Cells on the flanks of the meristem contribute to production of leaves. The adaxial-abaxial axis is marked. (b) Mature leaf showing the proximal-distal and medial-lateral axes.
The resulting basic leaf shape is a flat, planar structure but modifications during development result in variations on the basic shape. For example, other shapes include leaves that are flattened along a lateral dimension or are radial, such as tendrils of Pisum sativum (pea) leaves or spines in many cacti. Varying degrees of serrations and lobing at the leaf margins also contribute to differences in leaf shape. For instance, within Australian Banksia, B. grandis leaves have deeply indented margins, whereas B. integrifolia leaves have smooth margins (Figure 7.12).
7.1-Ch-Fig-7.12v2.jpg

Figure 7.12 Contrasting leaf shape of (left) Banksia grandis and (right) Banksi integrifolia. (Photographs courtesy M. Fagg and D. Greig, ANBG)
In addition to these types of elaborations, the leaf lamina may be a simple undivided shape or may be complex and subdivided into leaflets. Complex or compound leaves are found in many species including tomato (Solanum lycopersicum) and pea (Pisum sativum) (Figure 7.13).
7.1-Ch-Fig-7.13v2.png

Figure 7.13 Leaf shape variation. Banksia serrata leaf has serrated margins (A) and B. integrifolia leaf has smooth margins (B). The leaf of Arabidopsis thaliana is simple with small marginal serrations (C). The leaf of tomato is compound and a single leaf has primary and secondary leaflets (D). The leaf of pea is compound and includes central leaflets, proximal stipules and distal radial tendrils (E).
Different leaf shapes may be produced during the life of the plant, and may be influenced by environmental conditions. For instance many Eucalyptus species have short, broad juvenile leaves and narrow, elongated adult leaves. Leaves in Acacia species change with plant maturity where juvenile leaves have a dissected leaf lamina and adult leaves (which are a modified petiole, or phyllode), are simple in shape (Figure 7.14).
7.1-Ch-Fig-7.14.jpg

Figure 7.14 Branch of an Acacia species showing two leaf shapes on the one plant. The juvenile leaves are compound leaves and are subdivided into leaflets. The adult leaves (phyllodes) are simple in shape.
Leaf initiation
7.1-Ch-Fig-7.14.jpg

Figure 7.14 Branch of an Acacia species showing two leaf shapes on the one plant. The juvenile leaves are compound leaves and are subdivided into leaflets. The adult leaves (phyllodes) are simple in shape.
Leaves initiate in peripheral regions of the shoot meristem (Figure 7.15). Sites of initiation occur where the hormone auxin forms a localised maximum. Local high levels of auxin are achieved through the action of a membrane-localised auxin efflux carrier PIN-FORMED1 (PIN1). The distribution of PIN1 to the plasma membrane on just one side of cells leads to directed flux of auxin away from sites where organs have already initiated and toward the site where organs will next form (Figure 7.15). Coincident with high levels of auxin, class I homeodomain transcription factor KNOX genes are down-regulated in initiating organs. Class I KNOX genes are expressed in the shoot meristem and are essential to maintain growth of the shoot meristem (Figure 7.15).
7.1-Ch-Fig-7.15.jpg
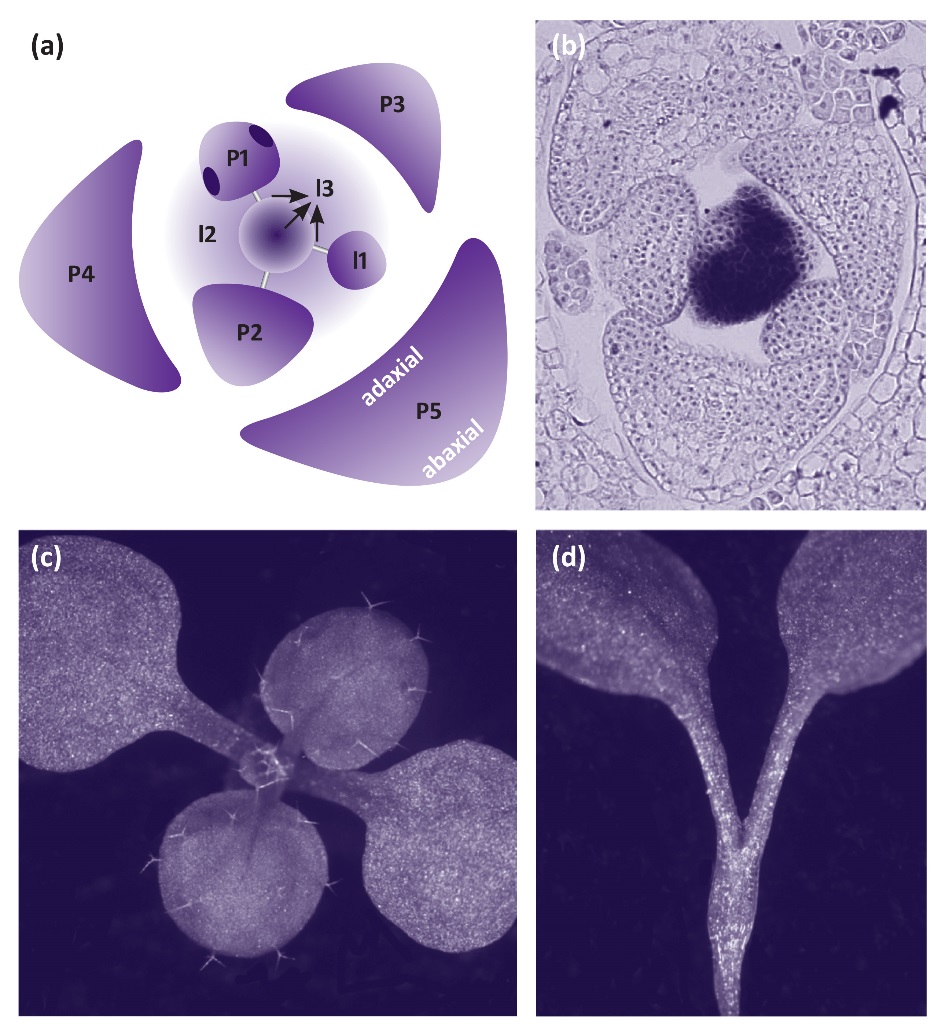
Figure 7.15 Leaf initiation and shape is determined by auxin and KNOX genes. (a) Diagram of the shoot apex with central meristem, incipient leaves I1-I3, and initiated leaves P1-P5. Arrows indicate the flow of auxin to the site where a new leaf will initiate. (b) in situ hybridisation showing expression of the class I KNOX gene SHOOT MERISTEMLESS in the shoot meristem and down-regulation in initiating and developing leaves. (c) Young Arabidopsis wild type seedling showing two cotyledons and first leaves. (d) The class I KNOX gene mutant shoot meristemless (stm) has 2 cotyledons and fails to produce leaves due to lack of a shoot meristem.
Each initiating leaf primordium is flanked by a boundary of slowly dividing cells and this boundary serves to delineate the leaf from the meristem and from adjacent leaf primordia. Several members of a transcription factor gene family known as CUP-SHAPED COTYLEDON (CUC) are expressed at this boundary and downregulation of CUC genes results in fusion of adjacent organs. This reflects the importance of the meristem-organ boundary in leaf development. As we see below, auxin, as well as the KNOX and CUC genes are also involved in determining leaf shape.
Proximal-distal axis
Following initiation leaf primordia go on to develop as discrete organs through cell division, cell expansion and cell differentiation. Leaf primordia become defined morphologically through cell divisions that lead to outgrowth from the meristem and hence formation of a proximal-distal axis. In the model species Arabidopsis thaliana (Arabidopsis), as the leaf grows a gradient of maturation occurs along this axis so that cells cease division and undergo terminal differentiation firstly in the distal region and lastly in the proximal region of the leaf. The leaf proximal-distal axis may have distinguishing features of lamina and petiole as in Arabidopsis and many dicotyledonous plants. One variation of this shape occurs in monocotyledons, which includes the grasses such as wheat, barley, rice and maize. Strap-like leaves of many monocotyledons have a distal blade and a proximal sheath, which wraps around the stem. The blade and sheath are separated by specialised cells that form a hinge, and this hinge allows the blade to bend away from the sheath and stem to be exposed to light.
Adaxial-abaxial axis
The adaxial-abaxial axis is evident at early stages of development of the leaf primordium. The adaxial side of the leaf is the side nearest to the shoot meristem and the abaxial side is furthest from the meristem (see Figure 7.11 above). Experimental separation of initiating leaf primordia from the shoot meristem results in loss of adaxial fate. Amazingly this loss of adaxial fate leads to development of an abaxial, radial leaf. This indicates that adaxial fate is determined either by contact with the meristem, or by signalling from the meristem.
Most flat leaves have adaxial-abaxial polarity where the two sides of a leaf have distinct morphological features. On the leaf surface, adaxial and abaxial epidermal cell size and shape, stomata density, or the presence of elaborated cell types such as hairs may be different. Vasculature often has a discrete adaxial-abaxial polar arrangement with adaxial water-conducting xylem and abaxial nutrient-conducting phloem (Figure 7.16a). Internal mesophyll cells of the leaf may also display differential arrangement along the adaxial-abaxial axis. In some species, the adaxial domain has closely aligned and chloroplast-rich palisade mesophyll cells whereas the abaxial domain has dispersed spongy mesophyll cells. Mature leaves are typically oriented with the adaxial side facing the light and the abaxial side away from the light. As such, distinct cell types along the adaxial-abaxial axis serve to optimize light capture and gas exchange for photosynthesis.
On top of this layer of regulation, adaxial and abaxial fates are each determined by specific genes. A reduction in expression of genes required for either adaxial or abaxial fate results in development of leaves that comprise only the opposite fate. Furthermore these leaves are radial. Together such observations have led to a fundamental tenet of leaf development, whereby adjacent adaxial and abaxial tissue types are essential to development of an adaxial-abaxial axis and hence to formation of a flattened leaf.
7.1-Ch-Fig-7.16.jpg
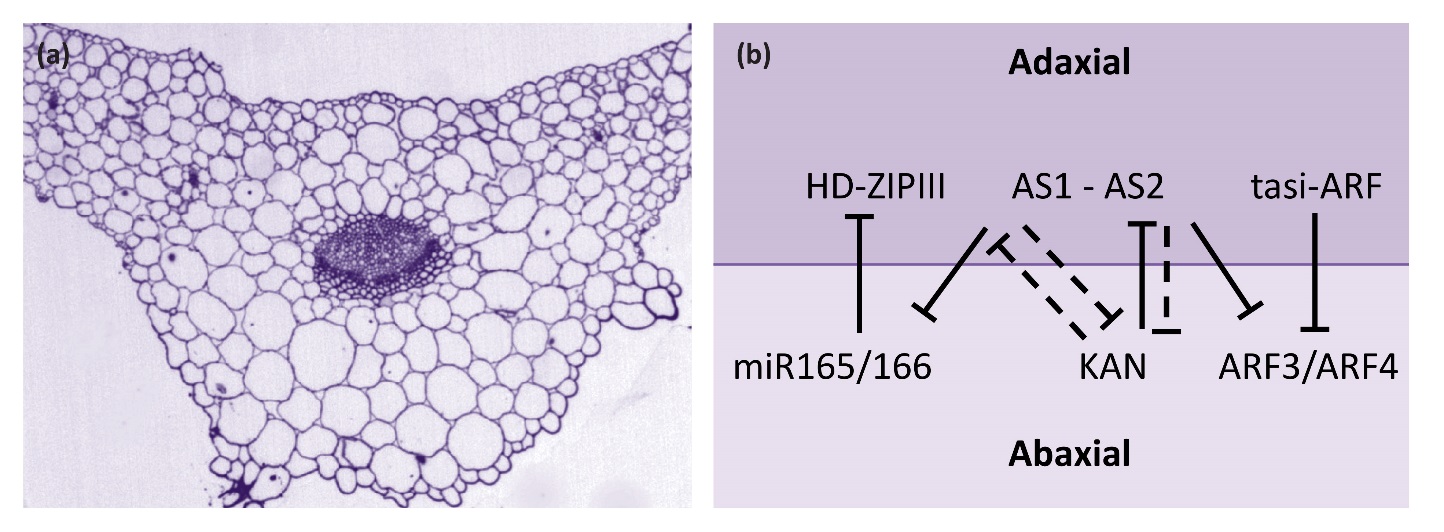
Figure 7.16 Leaf adaxial-abaxial fate. (a) Cross section through the mid-vein region of an Arabidopsis leaf. Tissues of the leaf including vasculature show distinct adaxial (upper surface) and abaxial (lower) differences. (b) Diagram showing adaxial and abaxial gene interactions. Solid lines indicate direct repression, dashed lines indicate indirect repression.
Key genetic determinants of adaxial fate include class III HD-ZIP transcription factors (Figure 7.16b). These proteins have a homeodomain, which is involved in binding DNA, a leucine zipper domain, which is involved in protein-protein interaction, and a sterol-binding domain that is predicted to bind small lipid molecules. In Arabidopsis, there are five genes encoding class III HD-ZIP transcription factors. Three of these genes, PHABULOSA (PHB), PHAVOLUTA (PHV) and REVOLUTA (REV), have a significant role in leaf development. Transcripts of all three genes are confined to the adaxial domain through transcriptional and post-transcriptional repression in the abaxial domain. For example, PHB, PHV and REV transcripts are all targets of the microRNAs miR165 and miR166, which are expressed on the abaxial side of the leaf. In mutants with reduced class III HD-ZIP activity adaxial fate diminishes. Conversely, gain of class III HD-ZIP expression in the abaxial domain results in conversion of the abaxial side of the leaf to adaxial fate.
Two other transcription factors, ASYMMETRIC LEAVES1 (AS1) and ASYMMETRIC LEAVES2 (AS2), form heterodimers and promote adaxial fate, in part through repression of miR165/166. Abaxial fate is promoted by additional factors, including KANADI (KAN) proteins, which are part of a plant-specific group of transcription factors. In Arabidopsis, four KAN genes have overlapping roles in leaf development. KANADI proteins are expressed on the abaxial side of the leaf and are necessary and sufficient for specifying the abaxial fate. It is also known that KAN1 directly represses AS2. Other abaxial factors are AUXIN RESPONSE FACTOR (ARF) proteins, which bind to promoter sequence elements in auxin responsive genes and regulate plant response to auxin. The two ARF genes, ARF3 and ARF4, are repressed on the adaxial side to the leaf by a set of small RNAs known as tasiR-ARF. The AS1-AS2 module directly represses ARF3. Thus the emerging picture is that repressive interactions between adaxial and abaxial genes helps to set up the adaxial-abaxial axis of the developing leaf (Figure 7.16).
Mediolateral axis
The mediolateral axis of leaves extends from the margin to the mid-region of the leaf, which is characterised by the presence of the main vein or midrib. Most leaves are bilaterally symmetrical and the two halves of the leaf, from margin to midrib, form mirror images. Leaf width is determined by the degree of lateral expansion along this axis. However, the relationship between the leaf margin and the midrib depends on cell recruitment from the shoot meristem during the process of initiation. Leaves of dicots initiate from a limited region of the shoot meristem so that the margins arise from cells close to the mid-region and lateral expansion of the leaf occurs post-initiation. On the other hand, leaves of monocots such as grasses, recruit cells from around the entire circumference of the shoot meristem. In this case, the leaf margins are derived from cells that are distant from the mid-region and consequently this early lateral expansion results in the leaf primordium wrapping around the entire shoot meristem.
Margins
Relatively simple modifications of the basic leaf shape are achieved through changes in growth dynamics at the leaf margins. In this way, leaves can develop marginal teeth or serrations or may be lobed. Leaf shape is more dramatically modified in compound leaves, where the blade is divided into reiterating units or leaflets. Multiple orders of subdivision occur when leaflets themselves are further subdivided into leaflets, for example in the compound leaves of tomato, which may have primary and secondary leaflets. Surprisingly, studies on the simple, serrated leaf of Arabidopsis, the lobed leaf of the Arabidopsis close relative Cardamine hirsuta, and the compound leaf of tomato have found that genes involved in setting up leaf initiation from the shoot meristem also function to modify the shape of the leaf lamina. The class I KNOX genes are important for modified leaf shape in many species. These genes are not expressed in the simple leaf of Arabidopsis but are expressed in the lobed leaf of Cardamine and the compound tomato leaf. Misexpression of class I KNOX genes in Arabidopsis leaves generates lobed leaves and increasing expression of these genes in Cardamine and tomato results in more lobing or more leaflets. Likewise, decreasing class I KNOX gene levels in Cardamine leads to a more simple leaf shape. Thus the degree of leaf complexity can depend on the expression of growth promoting class I KNOX genes. In addition to KNOX genes, auxin and CUC genes play central role in elaboration of leaves. In compound leaves of tomato and lobed leaves of Cardamine, high levels of auxin occurs at discrete sites along the developing leaf margin, where leaflets or lobes will form. Furthermore CUC gene expression marks the boundaries between initiating leaflets. At these sites CUC genes act to repress growth thereby promoting separation of leaflets. Altering the spatial distribution of auxin or CUC in the young developing leaf leads to reduced leaflet production. Less intricate elaborations of leaf shape, such as leaf serrations, also involve auxin and CUC genes. In developing leaves of Arabidopsis, the margins form periodic auxin maxima flanked by boundaries of CUC gene expression (Figure 7.17).
7.1-Ch-Fig-7.17.jpg
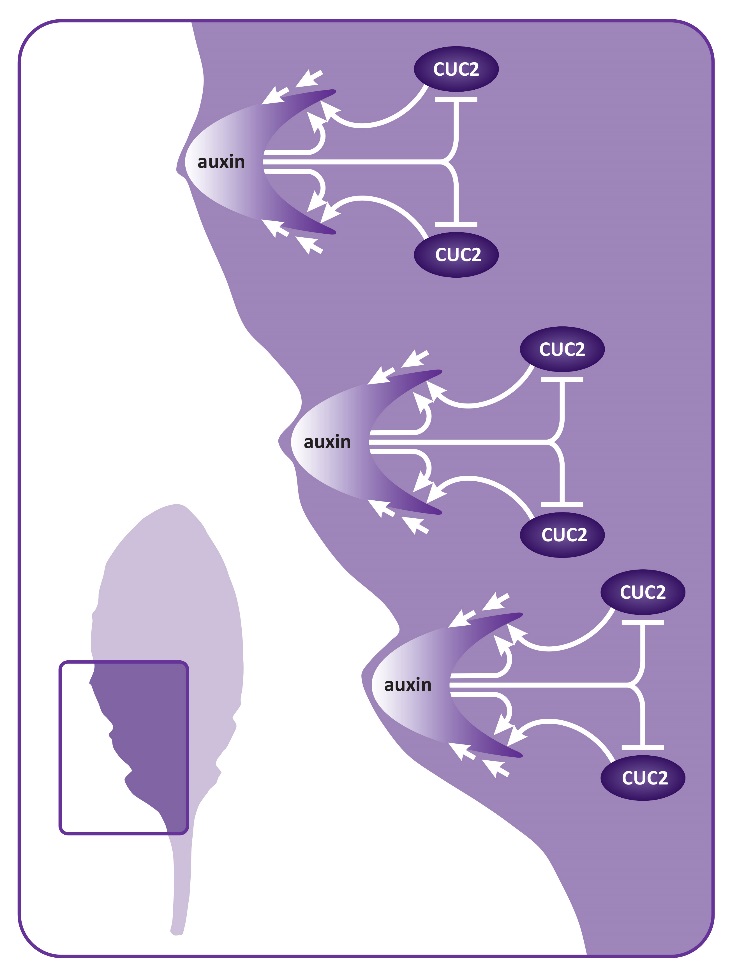
Figure 7.17 Modifying leaf margins. Serrations in Arabidopsis leaves occur at sites of auxin maxima. The PIN1 protein (straight arrows) promotes formation of an auxin peak. PIN1 is positively regulated by auxin and CUC2 (curved arrows). High levels of auxin in turn represses CUC2.
Further reading on leaf development
Bar M, Ori N (2015) Compound leaf development in model plant species. Curr Opin Plant Biol 23: 61-69
Bowman JL, Floyd SK (2008) Patterning and polarity in seed plant shoots. Ann Rev Plant Biol 59: 67-88
Braybrook SA, Kuhlemeier C (2010) How a plant builds leaves. Plant Cell 22: 1006-1018
Byrne ME (2005) Networks in leaf development. Curr Opin Plant Biol 8: 59-66
Byrne ME (2006) Shoot meristem function and leaf polarity: the role of class III HD-ZIP genes. PLoS Genet 2: e89
Chitwood DH, Nogueira FT, Howell MD et al. (2009) Pattern formation via small RNA mobility. Genes Dev 23: 549-554
Efroni I, Eshed Y, Lifschitz E (2010) Morphogenesis of simple and compound leaves: a critical review. Plant Cell 22: 1019-1032
Fukushima K, Hasebe M (2014) Adaxial-abaxial polarity: the developmental basis of leaf shape diversity. Genesis 52: 1-18
Hay A, Tsiantis M (2010) KNOX genes: versatile regulators of plant development and diversity. Develop 137: 3153-3165
Hepworth SR, Pautot VA (2015) Beyond the divide: Boundaries for patterning and stem cell regulation in plants. Front Plant Sci 6: 1052
Machida C, Nakagawa A, Kojima S et al. (2015) The complex of ASYMMETRIC LEAVES (AS) proteins plays a central role in antagonistic interactions of genes for leaf polarity specification in Arabidopsis. WIRES Develop Biol 4: 655-671
Mentink RA, Tsiantis M (2015) From limbs to leaves: common themes in evolutionary diversification of organ form. Front Genet 6: 284
Sluis A, Hake S (2015) Organogenesis in plants: initiation and elaboration of leaves. Trends Genet 31: 300-306.